Overview

- Tolkovsky, A. (1997) Trends Neurosci. 20, 1.
- Jing, S. et al. (1992) Neuron 9, 1067.
- Acheson, A. et al. (1995) Nature 374, 450.
- Morse, J.K. et al. (1993) J. Neurosci. 13, 4146.
- Hyman, C. et al. (1991) Nature 350, 230.
- Friedman, B. et al. (1995) J. Neurosci. 15, 1044.
- Meyer, M. et al. (1992) J. Cell Biol. 119, 45.
- Koliatsos, V.E. et al. (1993) Neuron 10, 359.
- Teng, H.K. et al. (2005) J. Neurosci. 25, 5455.
- Woo, N.H. et al. (2005) Nat. Neurosci. 8, 1069.
 Alomone Labs Recombinant mouse proBDNF protein mediates ERK1/2 activation in TrkB transfected HEK-293 cells.Three days post transfection, the cells were serum-starved for 2h and then challenged for 10 min with Recombinant human BDNF protein (#B-250) or Recombinant mouse proBDNF protein (#B-240). Cell proteins were resolved by SDS PAGE and the level of ERK1/2 phosphorylation determined using anti-phospho ERK.
Alomone Labs Recombinant mouse proBDNF protein mediates ERK1/2 activation in TrkB transfected HEK-293 cells.Three days post transfection, the cells were serum-starved for 2h and then challenged for 10 min with Recombinant human BDNF protein (#B-250) or Recombinant mouse proBDNF protein (#B-240). Cell proteins were resolved by SDS PAGE and the level of ERK1/2 phosphorylation determined using anti-phospho ERK.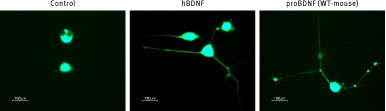 Alomone Labs Recombinant mouse proBDNF protein mediates neurite outgrowth in TrkB transfected PC12 cells.One day post-transfection, cells were stimulated with 0.2 nM Recombinant human BDNF protein (#B-250) or 0.2 nM Recombinant mouse proBDNF protein (#B-240). Development of neurites was visualized by fluorescence microscopy after 4 days.
Alomone Labs Recombinant mouse proBDNF protein mediates neurite outgrowth in TrkB transfected PC12 cells.One day post-transfection, cells were stimulated with 0.2 nM Recombinant human BDNF protein (#B-250) or 0.2 nM Recombinant mouse proBDNF protein (#B-240). Development of neurites was visualized by fluorescence microscopy after 4 days.
BDNF is a neurotrophic factor produced by proteolytic cleavage of its precursor, proBDNF. The biologically relevant form of the protein was thought to be the mature form, BDNF, which has been shown to affect the development of motoneurons, modulate synaptic transmission and affect axonal branching, dendrite growth and synapse number.1 These actions are mediated via the binding of BDNF to TrkB. Different actions are mediated by the binding of BDNF to p75 such as neuronal process retraction2 and neuronal apoptosis.3 The precursor form was thought to be important for the correct folding, secretion and trafficking of the mature protein. A single-nucleotide polymorphism (Val66 to Met) in the pro-domain of the human BDNF gene impairs intracellular trafficking and regulated secretion of BDNF in primary cortical neurons and neurosecretory cells but not in endothelial and vascular cells.4 This has been shown to affect memory and lead to abnormal hippocampal function in humans.5
The finding that proBDNF and not mature BDNF is the preferred ligand for p756 has ushered in a new era which reexamines the biological roles of the two forms. Contradictory biological roles for proBDNF have been proposed. It has been shown to be a pro-apoptotic ligand for sympathetic neurons7 expressing both p75 and sortlin, and to be involved in LTD8. On the other hand it has also been shown to elicit prototypical TrkB responses in biological assays, such as TrkB tyrosine phosphorylation, and activation of ERK1/2.9 Binding of both proBDNF and mature BDNF to TrkB has been proposed to be via the R103 residue in the mature portion.9 In addition, the question of the relative abundance of the precursor vs the mature form has been investigated. In brain homogenates a mixture of proBDNF and mature BDNF has been found.10,11 The secretion of proBDNF from cortical neurons has been shown.7 Most of the work showing secretion of proBDNF was done in cell lines overexpressing full length BDNF constructs.4, 6, 12, 13 It has been argued14 that this model misrepresents the authentic situation in vivo, since in the transfected systems the secretion of proBDNF results from overloading the limited capacity of the processing machinery. These authors argue that in fact proBDNF is a transient intermediate and is rapidly converted intracellularly to mature BDNF.
