GLP-1R agonists such as semaglutide (Ozempic), liraglutide (Victoza), and dulaglutide (Trulicity) are best known for enhancing insulin secretion and reducing glucagon levels in a glucose-dependent manner. These effects improve glycemic control without causing hypoglycemia, and they also delay gastric emptying and suppress appetite, contributing to weight loss. These actions reflect GLP-1R activity in pancreatic β-cells and gastrointestinal tissues.
The Multifaceted Role of GLP-1R
GLP-1R is now being studied for its immunomodulatory potential. Recent studies show that GLP-1R is expressed on CD4+ T cells and macrophages, where it shapes T cell differentiation and inflammatory tone (1–3). These findings show that GLP-1R signaling intersects with immune regulation pathways beyond metabolic control.
GLP-1R activation also influences T cell polarization and cytokine output. In vitro and in vivo studies have shown that GLP-1R agonists increase regulatory T cell frequency, reduce CD+ Th17 (T helper 17) cell differentiation, and dampen pro-inflammatory cytokines such as interleukin (IL)-6 and TNF-α (2, 4, 5). These effects are thought to be mediated through cAMP/PKA signaling downstream of the GLP-1R, leading to suppression of NF-κB activation and reduced STAT3 transcription factor phosphorylation. In mouse models, GLP-1R agonists have been shown to mitigate inflammation in contexts ranging from colitis to cardiovascular injury (5).
These immunoregulatory effects are drug-specific and context-dependent. For example, semaglutide and liraglutide have both been reported to increase IL-10 production in monocytes, while exenatide reduces macrophage infiltration in adipose tissue (6, 7). However, the precise distribution, trafficking, and surface accessibility of GLP-1R on immune cells remain incompletely characterized.
Despite the growing interest in this research area, these receptors have been difficult to study in live immune cells due to their low expression and poor antibody accessibility.
Alomone’s Extracellular GLP-1R Antibodies Support Live Cell Imaging and Flow Cytometry
At Alomone Labs we offer a polyclonal Anti-GLP1R (extracellular) Antibody (#AGR-021) to target the extracellular domain of GLP-1R (Figure 1). This antibody was developed and tested in-house, which allows for the labeling of live, non-permeabilized cells – important for studying receptor localization and trafficking in pancreatic islets and subsets of immune cells.
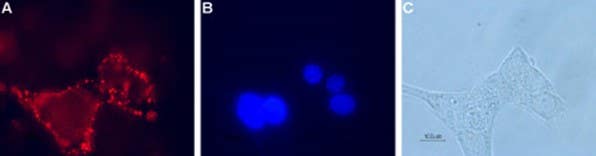
Figure 1. Expression of GLP-1R in RIN-5F, a rat insulinoma-derived pancreatic β-cell line. Cell surface detection of GLP-1R in live intact pancreatic islet cells. A) Cells were stained with Anti-GLP1R (extracellular) Antibody (#AGR-021), (1:100), followed by goat anti-rabbit-AlexaFluor-594 (red). B) Cell nuclei were visualized with the membrane-permeable DNA dye Hoechst 33342 (blue). C) Live view of the cells.
The antibody is also available conjugated to FITC (Anti-GLP1R (extracellular)-FITC Antibody (#AGR-021-F) or R-phycoerythrin (R-PE) (Anti-GLP1R (extracellular)-PE Antibody (#AGR-021-PE). These conjugated versions enable surface-level detection of GLP-1R on live immune cells by flow cytometry, which is not possible with intracellular antibodies or antibodies directed against denatured epitopes.
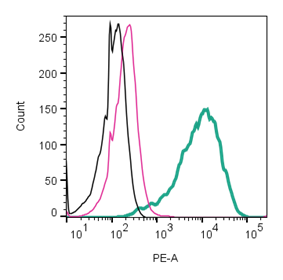
Figure 2. Cell surface detection of GLP-1R by direct flow cytometry in live, intact mouse J774 macrophage cells. Black) Untreated cells. Pink) Cells treated with Rabbit IgG isotype control-PE (#RIC-001-PE). Green) Cells stained with Anti-GLP1R (extracellular)-PE Antibody (#AGR-021-PE), (2.5µg).
Our antibodies allow for the tracking of receptor dynamics in real-time across diverse immune phenotypes. For example, GLP-1R was detected on live CD4+ T cells and pancreatic islets using our FITC-conjugated antibody (#AGR-021-F) for flow cytometry and immunocytochemistry. Therefore, this confirms the surface accessibility of both antibodies and their utility for dynamic immune profiling (8).
To date, Alomone Labs’ #AGR-021-F and #AGR-021-PE remain among the few extracellular GLP-1R antibodies commercially available for flow cytometry and live cell immune profiling.
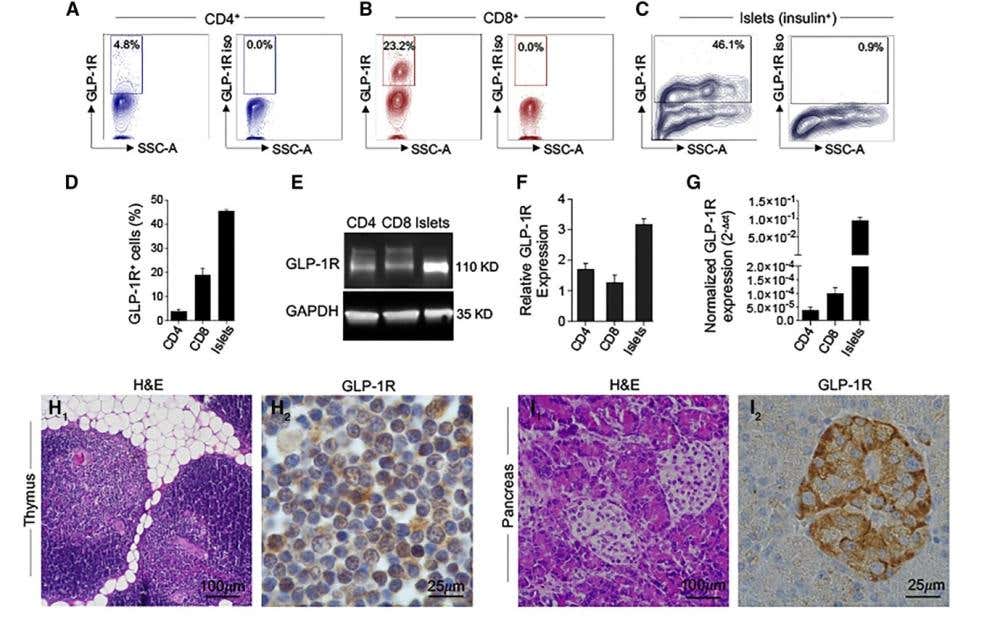
Figure 3. Expression and functional validation of GLP-1R in human T cells. A-C) Representative flow cytometry dot plots showing GLP-1R expression [y-axis: GLP-1R fluorescence intensity (GLP-1R); x-axis: side scatter area, (SSC-A)] in circulating CD4⁺ T cells (A), CD8⁺ T cells (B), and pancreatic islet cells [Islets (insulin⁺)], which were used as a positive control (C). D) Quantification of GLP-1R⁺ cell percentages of (A)-(C). E) Western blot analysis of GLP-1R (110 kDa) and GAPDH (loading control, 35 kDa) in CD4⁺ T cells, CD8⁺ T cells, and islets. F) Bar graph showing GLP-1R protein expression relative to the positive control (islets). Unstimulated CD3⁺ T cells were used as the negative control. G) Normalized GLP-1R mRNA expression (2⁻ΔΔCt) across the same cell types. H), I) Representative hematoxylin and eosin (H&E) and GLP-1R immunohistochemistry images in human thymus and pancreas, respectively. The data represent the mean ± SEM (n ≥ 3). *p < 0.05, **p < 0.01, ***p < 0.001. Adapted from Nasr et al., 2024 (8).
Validated Reagents to Track GLP-1R on Immune Cells and Islets In Vivo
Unlike intracellular antibodies or antibodies directed against denatured epitopes, surface-directed reagents detect physiologically accessible GLP-1R. This supports mechanistic studies into receptor trafficking, drug binding, and signaling – particularly in immunometabolic areas like pancreatic islets and inflamed adipose tissue.
While GLP-1R agonists are already established in metabolic disease, their expression in immune cells opens new avenues for mechanistic research. At Alomone Labs, we provide in-house developed and validated cell-surface-directed antibodies that support mechanistic studies of GLP-1R function in both metabolic and immune systems.
Explore our full catalog of antibodies targeting the glucagon receptor family, including tools for GLP-1R, Glucagon Receptor (GCGR), and related metabolic receptors.
 Antibody")
-FITC Antibody")