
When classic antibodies fall short in visualizing ion channels, you might want to consider nature’s own solution: toxins from animals like scorpion, spider, or cone snail venom, conveniently rendered visible by conjugating them to fluorescent probes.
Understanding the physiological and clinical significance of ion channels such as voltage-gated sodium (NaV) and potassium (KV) channels is at the core of a lot of neuroscience research. But mapping their precise distribution, especially in live tissues, poses a substantial technical challenge. While immunoassays are a standard method – using antibodies coupled to fluorescent tags or secondary antibodies (Table 1) – this approach comes with several comparative limitations: antibodies are large, can impact channel function, may require fixation (thus losing live context), and can have limited efficacy when targeting ion channel extracellular regions unless specifically designed to do so.
Why Use Toxins?
Natural peptide toxins have evolved to bind ion channels with exceptional selectivity, potency, and affinity, often at nanomolar or picomolar concentrations. Their small size and high specificity allow them to distinguish between even closely related channel subtypes. They can bind in living, unfixed tissue, bypassing the need for fixation and providing access to dynamic biological processes. Importantly, they offer the unique possibility to modulate channel function, serving as both tools for visualization and agents for mechanistic study.
These features have made toxins indispensable for functional studies in electrophysiology. Patch clamping, the gold standard for direct channel recording, uses toxins for their unique selectivity-but this offers no spatial information about channel distribution in the tissue.
This is where chemical ingenuity steps in. By linking a fluorophore directly to a toxin, you can create probes that retain the toxin’s precise targeting and gain the optical power of high-resolution microscopy. Examples include:
- Chimeric constructs such as fluorescent protein-scorpion toxin fusions, effective for visualizing KV channel subtypes in complex tissues (Kuzmenkov 2016).
- Bioconjugate combining chlorotoxin (CTX) with the far-red dye Cy5.5 enabled intraoperative imaging of cancer – including glioma, prostate, and intestinal tumors in mouse models. This “Tumor Paint” specifically illuminates malignant cells, offering potential high-resolution optical contrast in surgical settings (Veiseh 2007).
- Fluorescently labelled spider-venom peptides, such as μ-theraphotoxin-Tsp1a conjugated to near-infrared dyes (e.g., BODIPY, chlorin, IR800), have been applied to visualize NaV1.7 distribution in peripheral nerves. Tsp1a-Pra-IR800 showed strong fluorescent signals in peripheral nerves, excellent contrast in surgical models, and no detectable toxicity in mice or non-human primates – making it a promising tool for intraoperative nerve imaging and potential diagnostic use (Chow 2016).
As well as precise visualization of ion channel distributions and localizations in live cells and tissues, fluorophore-conjugated toxins also retain their native ability to modulate channel function.
This dual functionality gives you the power to simultaneously map channel spatial patterns and assess dynamic functional states. This is a truly integrated approach to studying ion channel physiology, making these powerful tools to dissect the relationship between channel localization and activity in physiological and pathological contexts.
Recent developments go even further. Some toxin-fluorophore probes now act as environment- and voltage-sensitive biosensors, providing not just location but also readouts of channel conformational state (Stewart 2021).
Table 1. Advantages of conjugated toxins over antibodies.
| Feature | Fluorescent Toxins | Antibodies |
| Size | Small (1–10kDa) | Large (~150kDa) |
| Live cell compatibility | Yes | Usually no |
| Selectivity/affinity | Extremely high | Variable |
| Target specificity | Excellent for subtypes | Sometimes broad |
| Perturbation of function | Possible (direct action) | Sometimes indirect |
| Multiplex/label flexibility | High | High |
Making Visuals with Venom Toxin
As you can see below, with a toxin conjugated with a fluorescent probe you can create beautiful and revealing microscopy images.

Figure 1. Live-cell imaging of Hm1a-ATTO Fluor-647N in differentiated PC-12 cells. Neurite outgrowth was induced by treating PC-12 cells with 100 ng/ml Native mouse NGF 2.5S protein (#N-240) for 10 days. A-B) Cellular membranes were labelled with CellMask™ Actin 1X solution (green). C-D) The same cells were then incubated with 0.2 µM Hm1a-ATTO Fluor-647N (#STH-600-FRN), producing red fluorescence at Hm1a binding sites. E-F) Overlay images highlight the distribution of Hm1a (red) relative to cell morphology (green) during live-cell imaging.
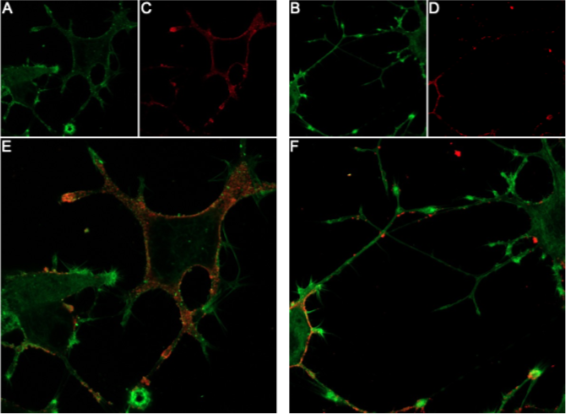
Figure 2. Live-cell imaging of GsMTx-4-ATTO Fluor-647N in differentiated PC-12 cells. PC-12 cells were differentiated by 10-day exposure to 100 ng/ml Native mouse NGF 2.5S protein (#N-240). A-B) Membranes were labelled with CellMask™ Actin 1X solution (green). C-D) Cells were then incubated with 0.1 µM GsMTx-4-ATTO Fluor-647N (#STG-100-FRN), producing red fluorescence at GsMTx-4 binding sites. E-F) Overlay images show the distribution of GsMTx-4 (red) across the membrane-labelled cells (green) during live-cell imaging.
Science with Sting
Fluorescently labelled toxins represent a powerful, technically sophisticated advance for live-cell and high-resolution ion channel research. By combining exquisite natural specificity with modern chemical and imaging technologies, these probes are propelling our understanding of channel function and distribution far beyond what traditional reagents allow.
If you’re curious about the practicalities – such as conjugation chemistries, validation protocols, or which toxin to use for your favorite ion channel – there’s a rapidly growing body of research and method articles to guide you.
Put a little more sting into your science and take a look at toxins


")

