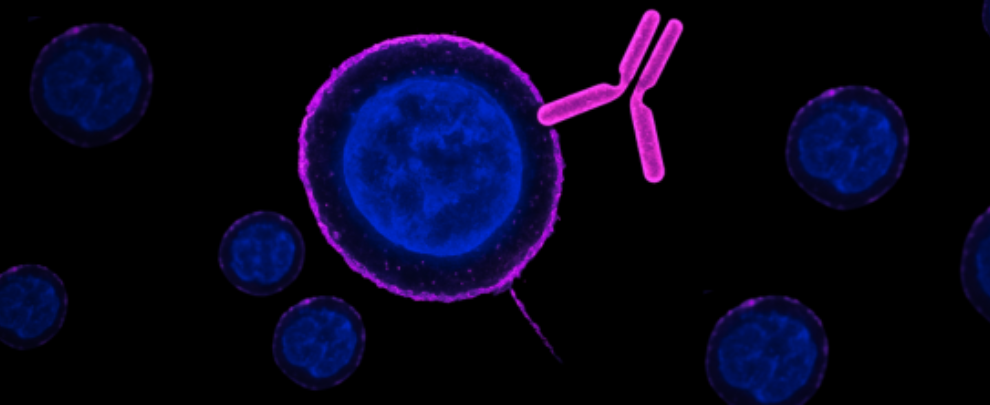
How cell surface-binding antibodies reveal real-time ion channel trafficking
There’s a whole world of biology happening at the cell surface. Ion channels shift positions. Receptors traffic in and out. Proteins translocate in response to stimuli. But most of the time, we only see the aftermath – because the tools we use require permeabilization or fixation. That means we’re always working from a snapshot of a system that’s already dead.
Cell surface-binding antibodies change that. They recognize extracellular epitopes – the domains of membrane proteins that stick out into the external environment. That one detail changes what you can do: now you can label surface proteins in live, intact cells, without disrupting their native state.
And that opens up the door to experiments that track protein movement in real time, with the kind of temporal and spatial resolution that fixed-cell approaches just can’t touch.
(In the past, we’ve always referred to these as “extracellular antibodies” because they bind the extracellular region of the protein. But it’s just not a very intuitive name, so we’ve decided to change it. From now on, we’ll call these cell surface-binding antibodies. We think it’s much clearer about what these antibodies actually do. Okay, enough housekeeping, let’s get back to the science!)
A Live-Cell Method That Works When the Cell Is Still Working
The key advantage here is that these antibodies bind before fixation – not after. You apply the primary antibody to live neurons, then fix the cells only after surface labeling has occurred. That’s exactly the approach used by Haan and Fisher in a pair of studies exploring how TRPV1 channels move to and from the cell membrane in response to osmotic stress (1, 2).
Using our Anti-TRPV1 (VR1) (extracellular) Antibody (#ACC-029), they captured a dynamic, reversible translocation process – one that only became visible because they were able to label the external face of the protein in real time, before fixation.
This isn’t just a niche protocol. It’s a broadly applicable way to visualize:
- Channel recruitment to the membrane
- Internalization after signaling
- Receptor cycling in response to physiological or pharmacological cues
You get real dynamics, not static structure. And you don’t need to genetically tag your protein of interest or overexpress anything.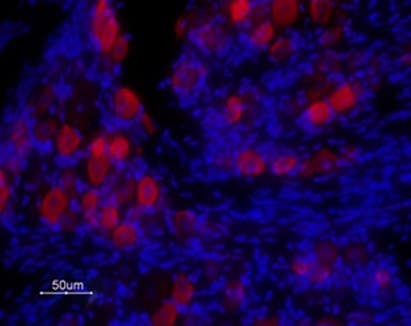
Figure 1. Alomone data (not from studies) showing expression of TRPV1 in rat DRG. Immunohistochemical staining of rat DRG frozen sections using Anti-TRPV1 (VR1) (extracellular) Antibody (#ACC-029, 1:100) followed by goat anti-rabbit-ATTO Fluor-594 secondary antibody (red). TRPV1 is expressed in medium and small DRG neurons. Hoechst 33342 is used as the counterstain (blue).
Here’s What This Looks Like in Practice
TRPV1 channels translocate to the surface during osmotic stress
When hypothalamic vasopressin neurons are exposed to high extracellular osmolality, TRPV1 channels move from intracellular stores to the plasma membrane. This increases the cell’s firing rate and promotes hormone release. It’s a slow, regulated process that happens over about 90 minutes – and it’s fully reversible when cells return to isotonic conditions.
How do we know this? Because the authors used live-cell immunocytochemistry with a cell surface-binding antibody to label only the external TRPV1 domains. That allowed them to isolate membrane-localized signal from total expression, and to watch the signal fade and reappear with changes in osmolality (1).
The same method reveals how that trafficking is controlled
In follow-up work, they laid out a full protocol for applying this approach in acutely isolated rodent neurons – including precise solution prep, oxygenation, pipetting, and antibody application. They showed how this method can be adapted to visualize the translocation of other surface proteins, and included a troubleshooting guide for ensuring clean membrane-bound signal (2).
Inhibiting exocytosis, PLC activity, or calcium influx blocked the TRPV1 translocation. That means this method doesn’t just tell you that trafficking occurs – it lets you test what drives it.
Why It Matters That the Antibody Binds the Outside
This is the critical feature. Because the antibody binds an extracellular epitope:
- You don’t have to permeabilize the membrane
- You preserve native cellular physiology during labeling
- You can distinguish surface-localized protein from internal pools
- You can capture trafficking as it happens
If the epitope were intracellular, you’d need detergent – and the whole point of watching live dynamics would be lost.
This is exactly why our cell surface-binding antibodies are validated for live-cell use. The anti-TRPV1 antibody used in these papers (#ACC-029) is specific to an extracellular epitope and we’ve tested it in both live and fixed conditions.
Want to Do This Yourself? Here’s What to Look for
If you’re planning to visualize membrane trafficking in real time, make sure your antibody is:
- Raised against an extracellular epitope
- Validated in live-cell or live-tissue conditions
- Compatible with the downstream method you plan to use (e.g., immunocytochemistry, flow, conjugation)
Then you can pair it with quantum dots, fluorescent secondaries, or cargo-carrying conjugates – and you’re ready to track how your surface protein behaves in the real, functioning cell.
Up Next in the Series
In Part 2 of our cell surface-binding antibodies, we’ll look at using these antibodies as delivery agents that enable targeted optoporation and cargo entry. Get ready for gold particles, femtosecond lasers, and the precise opening of the membrane – all driven by surface protein recognition.
 (extracellular) Antibody")