What happens when you follow receptors and ion channels without disrupting the system they live in? Cell surface-binding antibodies – designed to target extracellular domains – let you observe proteins in live tissue, track disease mechanisms in real time, and dissect signaling events as they happen.
In the previous part of this series, we showed how these antibodies allow real-time tracking (Part 1), targeted photomodulation (Part 2), and single-particle receptor dynamics (Part 3). In this final entry, we leave the culture dish behind and head into two complex biological systems: the memory-forming cortex and the invasive tumor microenvironment. Both studies highlight how extracellular antibodies (and related tools) let us study live, dynamic processes – without crossing the membrane or killing the cell.
Synapses in Motion – Tracking Receptors Across Brain Networks
Memory doesn’t live in one place. As engrams consolidate, cortical networks take over from the hippocampus. But what allows those remote circuits to stabilize – or stay flexible – hasn’t been clear. Bessières and colleagues tracked this directly in rats by using antibodies against NMDA receptor subunits, Anti-NMDA2R (GluN2A) (extracellular) antibody (#AGC-002) and Anti-NMDAR2B (GluN2B) (extracellular) antibody (#AGC-003) in combination with in vivo imaging and receptor-specific perturbations (1).
They showed that GluN2B-containing receptors are recruited early to encode memories, while GluN2A gradually takes over during stabilization. By using surface-binding antibodies to map receptor localization and behavior, they could observe a redistribution of NMDA receptor subtypes at synapses over time. The key was live tracking of extracellular epitopes – no need to overexpress tagged proteins or fix tissue.
When GluN2B subunits were pharmacologically retained at the synapse (instead of being replaced by GluN2A), memory engrams became unstable. Conversely, shifting receptor balance altered memory malleability (Figure 1). This would have been impossible to resolve with standard fixed-tissue staining.
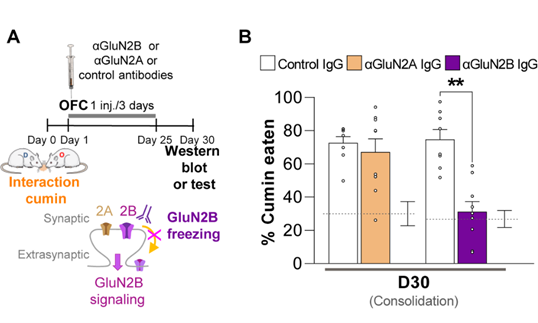
Figure 1. Targeting GluN2B in the Cortex Reshapes Remote Memory Storage. This figure is adapted from Bessières et al., Science Advances (2024) 10.1126/sciadv.ado1148. A) Experimental timeline illustrating the effect of antibody injections into the orbitofrontal cortex (OFC) on memory consolidation. Rats underwent a social interaction session with access to cumin-flavored food on Day 0, forming an odor-associated memory. Starting on Day 1, they received intracortical injections every three days with either anti-GluN2B (#AGC-003), anti-GluN2A (#AGC-002), or control IgG antibodies. The schematic highlights the targeting of synaptic and extrasynaptic NMDA receptors: GluN2A-containing receptors (2A) remain unaffected, while GluN2B-containing receptors (2B) are blocked by the antibody, reducing GluN2B signaling. Memory retention and protein levels were assessed around Day 30 via behavioral testing or Western blotting. B) Rats that had received the anti-GluN2B antibody showed significantly reduced consumption of cumin-flavored food after 30 days, indicating impaired recall of the original memory. This effect was not observed with the anti-GluN2A or control antibodies.
Visualizing Mechanotransduction and Ion Channel Trafficking in Tumor Invasion
To understand how ductal carcinoma in situ (DCIS) becomes invasive, Bu et al. used cell surface-binding antibodies to study TRPV4 channel localization in multiple physiologically relevant cancer cell lines (MCF10DCIS.com, MCF10CA1a, ETCC-006, ETCC-010, MDA-MB-231) (2). They found that under crowding stress, these otherwise contained cells could rapidly adopt an invasive phenotype.
Central to this shift was TRPV4, a stretch-activated calcium channel. By tagging TRPV4 with Anti-TRPV4 (extracellular) antibody (#ACC-124), the team followed its membrane recruitment in live tissue and quantified downstream changes in calcium influx, cell volume, and invasiveness. Under crowding stress, it moved from internal compartments to the membrane but remained inactive. This suppressed calcium entry, reduced cell volume, and triggered a pro-invasive phenotype.
But TRPV4 wasn’t acting alone. Two other channels – KCa3.1 (KCNN4) and Piezo1 – also relocalized to the surface during crowding and contributed to the invasive response. The researchers made use of Anti-KCNN4 (KCa3.1, SK4) (extracellular) antibody (#ALM-051) and Anti-Piezo1 (extracellular) antibody (#APC-087) to get the full picture.
Together, these antibodies allow precise detection of surface protein shifts under physiological stress. Functionally, blocking TRPV4 or disrupting KCNN4 and Piezo1 altered cell volume and reduced invasive behavior – proving their role in the biomechanical pathway. Without these cell surface-binding antibodies, the rapid relocalization of each channel would’ve been difficult to capture.
By targeting the extracellular domain, the team avoided the need to permeabilize cells, ensuring they could track native localization patterns as cells responded dynamically to their microenvironment.
Why It Matters – and What We’ve Seen
Across this series, we’ve explored what happens when you target the outside of the cell – and leave everything else intact. In Part 1, we tracked how surface-binding antibodies reveal the hidden movement of ion channels in real time, without permeabilization or genetic modification. In Part 2, we saw how those same antibodies could anchor cargo to the membrane – allowing researchers to deliver materials or trigger cellular events with pinpoint precision using light and gold nanoparticles. In Part 3, we looked at quantum dots and single-particle tracking to watch receptor dynamics at nanometer scales on living neurons. Now in Part 4, we’ve seen these antibodies taken into complex live systems.
The value of these cell surface-binding antibodies comes from what they don’t do: they don’t cross membranes. They don’t require you to kill your cells. They don’t rely on overexpressing or mutating proteins. Instead, they bind cleanly and specifically to what’s already there, making dynamic processes visible – without disrupting them.
The outside of the cell is where biology meets the world. These antibodies help us watch what happens next.
 (extracellular) Antibody")
 (extracellular) Antibody")
 Antibody")
 (extracellular) Antibody")
 Antibody")