α-Bungarotoxin (α-BgTx) is a 74 amino acid peptidyl toxin isolated from the venom of the banded krait snake, Bungarus multicinctus (1).
It blocks postsynaptic neuromuscular transmission via competitive inhibition of nicotinic ACh receptors (nAChRs) with an IC50 of 3.5 x 10-10 M, thereby prevents the depolarizing action on postsynaptic membranes and blocks neuromuscular transmission (2).
The toxin is selective for nAChR α7 receptors (IC50 value of 1.6 nM) and nAChR α3/β4 receptors (IC50 value of > 3 μM) (3, 4). α-BgTx also binds to, and blocks, a subset of GABAA receptors (GABAARs) that contain the GABAAR β3 subunit. In particular, α-BgTx blocks GABAARs that contain interfaces between adjacent β3 subunits (5).
Beyond its established role in labeling and studying nicotinic acetylcholine receptors, α-BgTx has become a valuable tool in modern research applications, including high-throughput drug screening and neurotoxicity studies. In drug discovery, α-BgTx’s high selectivity for α7 nAChRs makes it ideal for screening candidate compounds that modulate cholinergic signaling, particularly in the context of neurodegenerative disorders and cognitive enhancement therapies (6). In neurotoxicology, α-BgTx is used to monitor receptor integrity and function, enabling sensitive detection of toxin- or drug-induced synaptic damage (7).
Recent studies have demonstrated the value of fluorescently labeled α-Bungarotoxin (α-BgTx) as a sensitive tool in neurotoxicology for assessing receptor integrity at the neuromuscular junction (NMJ). Ionescu et al. employed FITC-conjugated α-BgTx to label nicotinic acetylcholine receptors (nAChRs) in a mouse model of amyotrophic lateral sclerosis (ALS). This method enabled precise visualization of disease-induced synaptic damage and facilitated evaluation of the neuroprotective small molecule pridopidine, which significantly restored NMJ structural integrity (8). These findings underscore the value of labeled α-BgTx in both mechanistic studies of neurotoxicity and preclinical drug development.
We are pleased to offer a comprehensive panel of α-Bungarotoxin conjugates and native toxin for a wide range of applications, including fluorescence imaging, receptor binding studies, and more. Our offerings include:
- α-Bungarotoxin (#B-100) – Native toxin, ideal for receptor binding and inhibition studies.
- α-Bungarotoxin-FITC (#B-100-F) – FITC-labeled conjugate, widely used for standard fluorescence assays.
- α-Bungarotoxin-Biotin (#B-100-B) – Biotinylated form, perfect for streptavidin-based detection.
We offer a set of ATTO-fluorophore–conjugated α-BgTx, as well, specifically designed for advanced imaging. These high-performance dyes are especially suited for super-resolution microscopy techniques, such as SIM and STED, and deliver exceptional signal quality:
- α-Bungarotoxin-ATTO Fluor-488 (#B-100-AG) – Labeled with green fluorescent ATTO Fluor-488 (Figure 1).
- α-Bungarotoxin-ATTO Fluor-550 (#B-100-AY) – Labeled with orange-red ATTO Fluor-550 (Figure 2).
- α-Bungarotoxin-ATTO Fluor-590 (#B-100-AR) – Provides red fluorescence.
- α-Bungarotoxin-ATTO Fluor-633 (#B-100-FR) – Offers deep red emission.
- α-Bungarotoxin-ATTO Fluor-647N (#B-100-FRN) – Far-red fluorophore with (Figure 3).
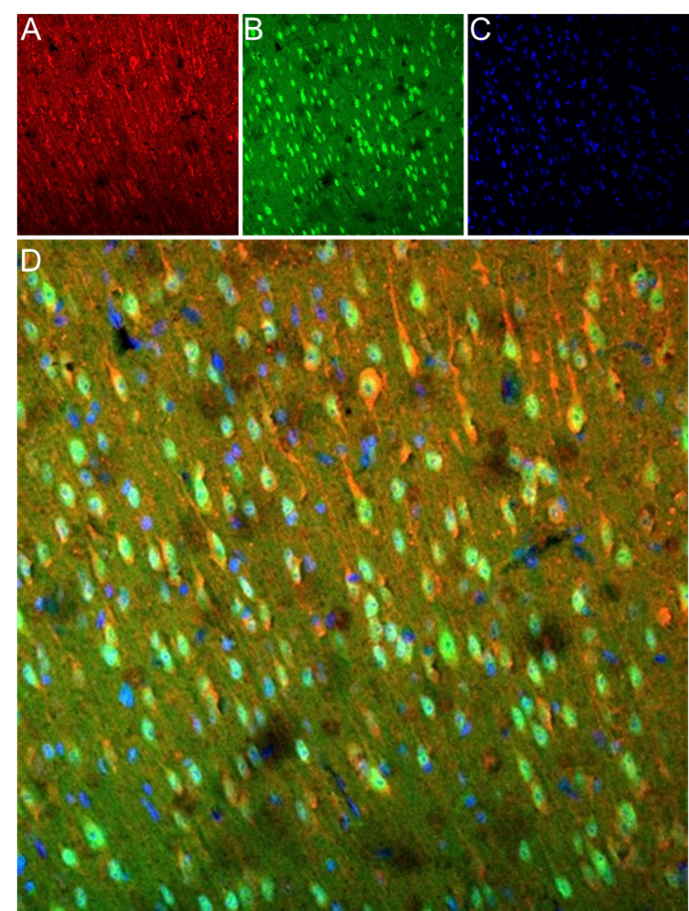
Figure 1. Expression of α-Bungarotoxin-ATTO Fluor-488 in mouse brain frozen sections. A) CHRNA7 (#ANC-007) was applied for 60 minutes at 37ºC, 1:100, followed by staining with goat anti-rabbit AlexaFluor-647, resulting in red fluorescence predominantly in cortex pyramidal neurons. B) The same frozen sections underwent incubation with 0.8 µM of α-Bungarotoxin-ATTO Fluor-488 (#B-100-AG) for 60 minutes at 37ºC, resulting in green fluorescence in neuronal cells. C) Cell nuclei were counterstained with Hoechst 33342, emitting blue fluorescence. D) Co-localization of CHRNA7 and α-Bungarotoxin-ATTO Fluor-488 appears in some cortex pyramidal neurons (indicated by arrows).
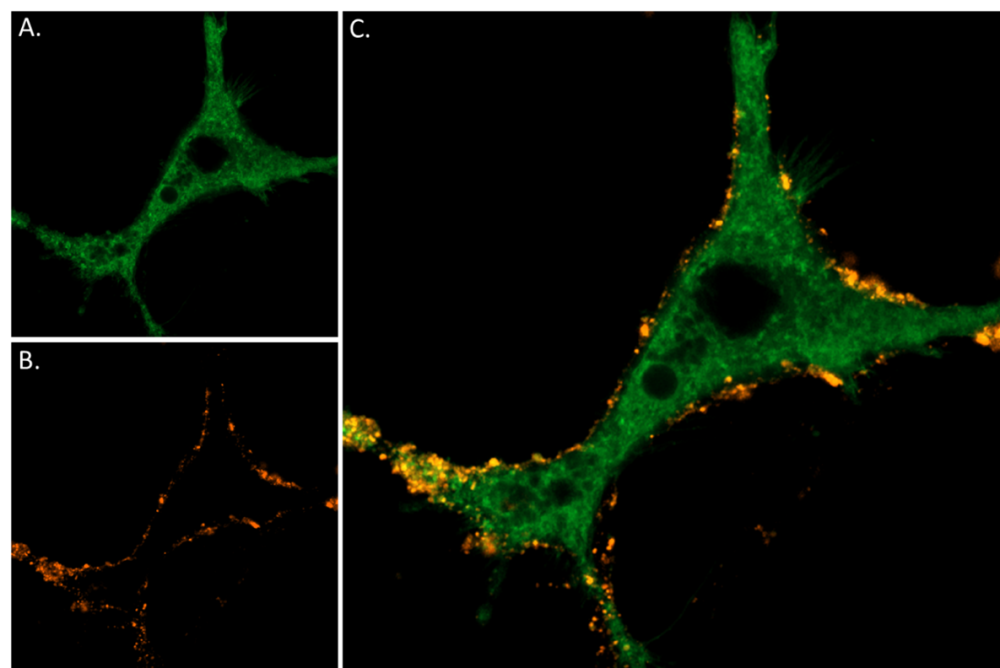
Figure 2. Live cell imaging of α-Bungarotoxin-ATTO Fluor-550 in differentiated PC-12 cells. Neurite outgrowth was induced in PC12 cells through 7 days exposure to 100 ng/ml Native mouse NGF 2.5S protein (99%) (#N-240). A) CellMask™ Actin 1X solution was applied for 30 minutes, resulting in a green fluorescence to visualize cellular membrane. B) Following this, the same cells underwent incubation with 0.3 µM of α-Bungarotoxin-ATTO Fluor-550 (B-100-AY) for 60 minutes at 37ºC, followed by PBSX1 wash, leading to orange (seen in red) fluorescence indicative of the distribution of nicotinic ACh channels. C) Merged image of panels A and B, showing the co-localization of the cellular membrane (green) and nicotinic ACh receptor distribution (red) in differentiated PC12 cells.
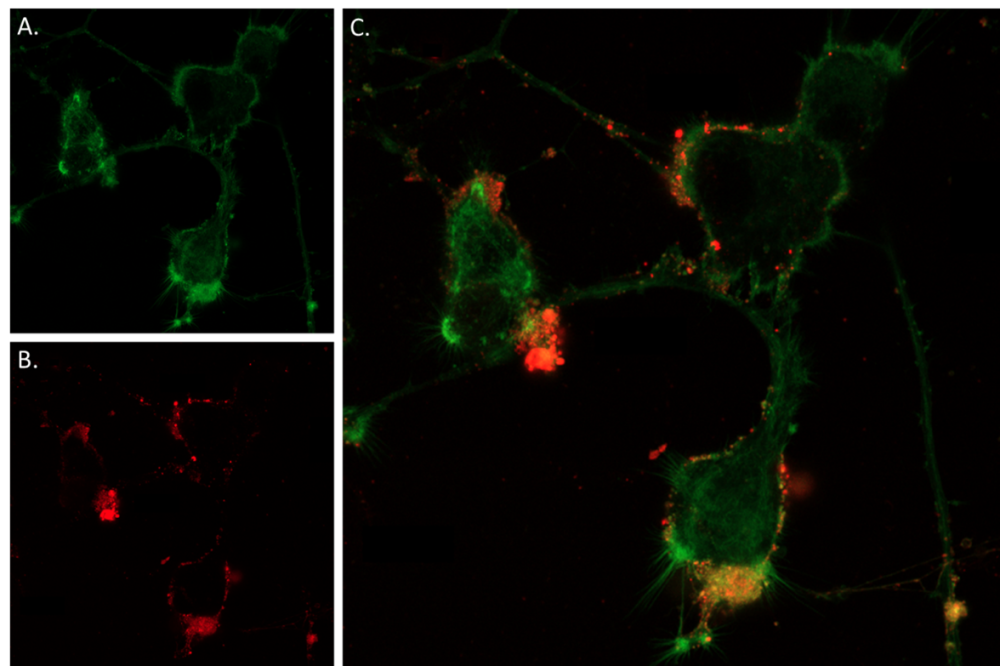
Figure 3. Live cell imaging of α-Bungarotoxin-ATTO Fluor-647N in differentiated PC-12 cells. Neurite outgrowth was induced in PC12 cells through 8 days exposure to 100 ng/ml Native mouse NGF 2.5S protein (99%) (#N-240). A) CellMask™ Actin 1X solution was applied for 30 minutes, resulting in a green fluorescence to visualize cellular membrane. B) Following this, the same cells underwent incubation with 0.1 µM of α-Bungarotoxin-ATTO Fluor-647N (B-100-FRN) for 60 minutes at 37ºC, followed by PBSX1 wash, leading to red fluorescence indicative of the distribution of nicotinic ACh channels. C) Merged image of panels A and B, showing the co-localization of the cellular membrane (green) and nicotinic ACh receptor distribution (red) in differentiated PC-12 cells.
Another interesting application is the use of the Bungarotoxin Binding Site (BBS) as a versatile molecular tag, enabling precise, real-time visualization of membrane proteins in live-cell environments. By engineering the short BBS peptide sequence into an extracellular domain of a protein of interest, researchers can detect specific, high-affinity interactions using labeled α-bungarotoxin in a non-invasive way. This strategy is particularly valuable for studying protein trafficking, turnover, and localization under physiological conditions, while avoiding the steric hindrance sometimes caused by larger fluorescent protein tags. When combined with advanced imaging tools, BBS facilitates multiplex experiments, tracking of receptor dynamics, and quantitative analyses of protein–ligand interactions. For more on the method’s benefits, see advantages of the Bungarotoxin Binding Site for studying live membrane protein dynamics and live-cell imaging of AMPA receptors using BBS and α-bungarotoxin.
These applications are further strengthened by advances in imaging and assay platforms, allowing real-time, quantitative analysis of receptor–ligand interactions (9).