Hearing when sound happens.
From the crack of a twig to the roll of a vowel, your brain decodes sound timing with millisecond precision. A growing body of research has suggested that this technical feat of temporal coding is the driving force behind how we perceive, protect, and preserve our sense of hearing. And when that timing fails, the whole auditory system can unravel.
Unlike vision, which can tolerate lag and blends signals over time, hearing depends on relentless temporal precision. Every word, footstep, or birdcall arrives not just with a pitch or loudness, but at a precise moment – and the brain relies on that timing to make sense of sound. Timing is the scaffold for everything the system does.
Four new studies have highlighted just how deeply hearing relies on this idea of timing: how neurons sharpen it, how circuits stretch it, how gene therapy can restore it, and what happens when it disappears.
Voltage-gated Potassium (KV)4 Channels Sharpen Temporal Precision at the First Auditory Relay
How do we achieve sharp timing from noisy input? Buried in the brainstem’s upper medulla is the ventral cochlear nucleus (VCN), where spherical bushy cells preserve the timing of auditory nerve inputs with sub-millisecond precision. A team from the US described the contribution of A-type potassium currents (primarily mediated by KV4.3 channels) to this process in mouse bushy neurons using Jingzhaotoxin-X (JZ-X, #STJ-080) or Phrixotoxin-1 (#STP-700), which are KV4-selective blockers (1).
KV4 currents were detected in 88% of bushy neurons. JZ-X blockade broadened action potential half-widths, reduced maximum rise slope, and increased spike latency and jitter during phase-locked auditory nerve stimulation (5–100 Hz). KV4 currents rapidly repolarized the membrane, shortened spike duration, and sharpened onset timing (Figure 1). Without the fast and transient A-type current coming from KV channels, neurons fired less precisely, degrading temporal fidelity, which is required for sound localization and speech discrimination. In vivo, this kind of timing jitter would likely disrupt the brain’s ability to compare sound arrival times between ears – a key mechanism for locating where sounds come from. Precision isn’t optional here – these neurons set the timing clock for everything that follows.
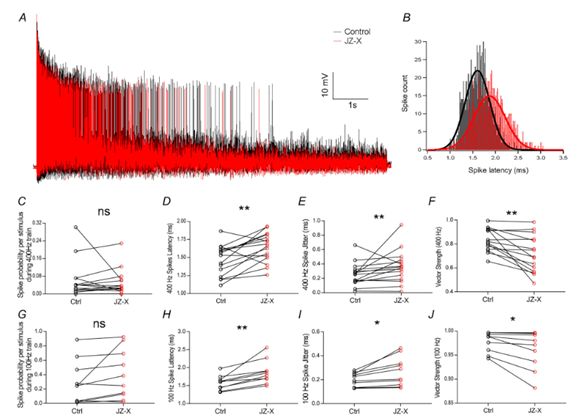
Figure 1. Jingzhaotoxin-X (JZ-X) disrupts the precise timing of neuronal firing. This figure shows how treatment with JZ-X affects the timing of electrical spikes (action potentials) in bushy neurons – cells that help process sound signals in the brain. A) The representative traces show neuron activity during 10-sec long rapid (400 Hz) stimulation of the auditory nerve: black for control and red after JZ-X treatment. B) The histogram summarizes when spikes occur relative to the stimulus. After JZ-X treatment (red), the spikes are delayed and more spread out compared to the control (black). C-J) Quantitative analysis showed that JZ-X didn’t alter how often the neurons fired compared to the control (Ctrl; C,G), but it did make them respond later (D, H: increased latency) and with less consistency (E, I: jitter). This indicated that their responses became less synchronized with the stimulus (F, J: reduced vector strength), both at 400 Hz (C-F) and 100 Hz (G-J). Statistical significance: ns = not significant; P < 0.05 (*), P < 0.01 (**). Image adapted from Zhang et al. (2024).
Medial Olivocochlear (MOC) Inhibition Slows Timing to Avoid Signal Overload
In contrast to afferent timing refinement, new studies showed how inhibition in the MOC system deliberately delays timing. MOC neurons provide feedback from the brainstem to the cochlea, to modulate outer hair cell activity (2). Even though the inhibitory ability is important for auditory function, the over-suppression of real signals needs to be avoided.
Using a “wedge-slice” preparation that preserved the full ascending auditory circuit, they stimulated either the auditory nerve (AN) or the medial division of the lateral lemniscus (MdL) while recording from identified MOC neurons. To isolate synaptic events, sodium channels were blocked with tetrodotoxin (TTX; #T-550) to ensure that recorded excitatory and inhibitory postsynaptic currents (EPSCs and IPSCs, respectively) reflected monosynaptic transmission rather than propagated spikes.
With MdL stimulation, excitation always preceded inhibition, indicating a polysynaptic inhibitory route. However, with AN stimulation, EPSCs and IPSCs arrived almost simultaneously, with similar latency and jitter. Machine learning classification of post synaptic currents (PSCs) combined with computational modeling (Figure 2) confirmed that this inhibitory-excitatory delay desynchronizes MOC output without abolishing it.
The result is a circuit that responds robustly to sustained background noise but ignores brief transient noise. The brainstem uses mistiming as a filter: MOC inhibition stretches the timing window to avoid maladaptive suppression of the cochlea. Across the auditory pathway, timing is tuned to purpose: where bushy cells chase precision, the MOC system slows it down, using fast inhibition to craft a slower, steadier response that preserves signal detection in noise.
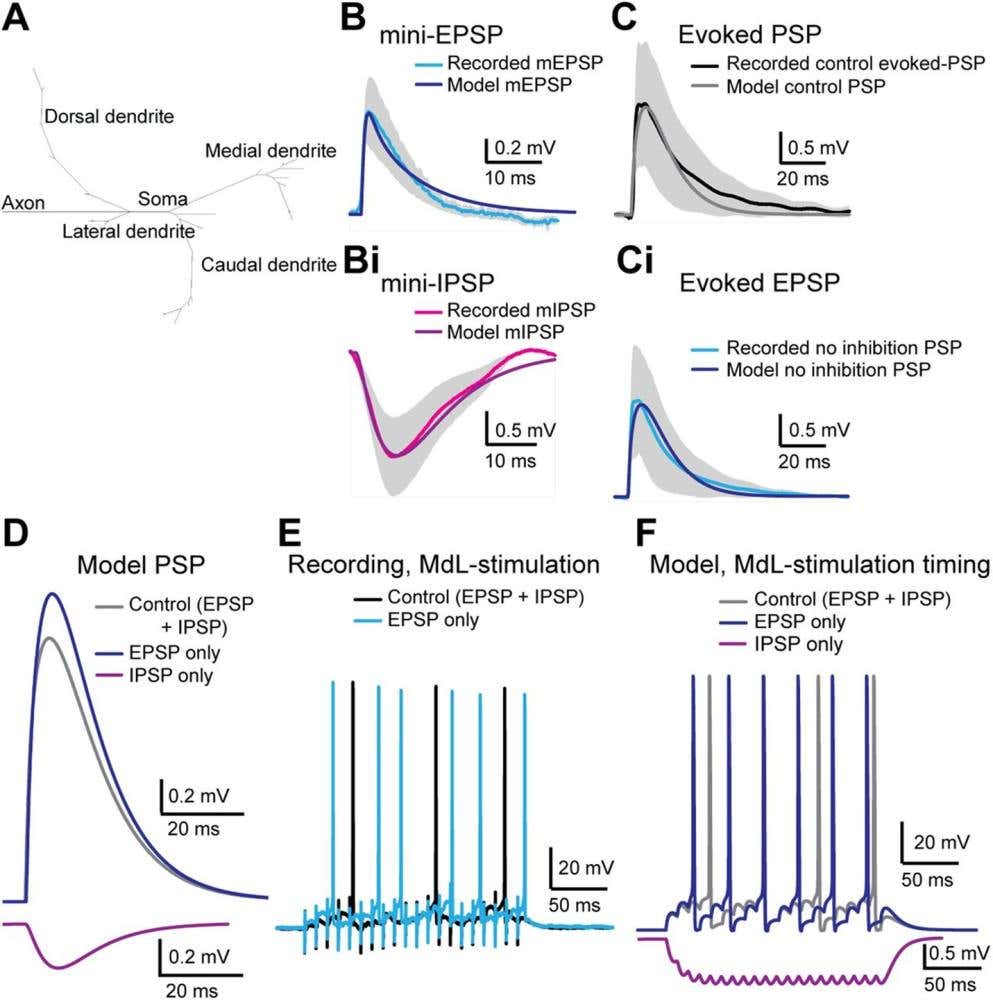
Figure 2. Modeling how inhibitory and excitatory signals shape medial olivocochlear (MOC) neuron responses. This figure compares real recordings from MOC neurons with a computational model designed to reproduce their electrical behavior. A) The model neuron structure includes dendrites (branches), a soma, and an axon. B-Bi) The miniature (m = mini) postsynaptic potential (PSP) was recorded from neurons – excitatory PSP (mEPSP = mini-EPSP, blue) and inhibitory PSP (mIPSP = mini-IPSP, magenta). The measured values closely match the model’s simulated responses (dark blue and purple, respectively). C-Ci) When larger evoked responses are tested, the model again mimics both the normal mixed excitation–inhibition response (black/gray) and the condition where inhibition is blocked, showing only excitation (blue). D) The model shows how excitatory and inhibitory inputs interact — with each shaping the total postsynaptic potential differently. E-F) Current-clamp recordings from MOC neurons during 100 Hz, 20 pulse medial olivocochlear (MdL) stimulation. Both real neurons (E) and the model (F) produce rhythmic firing patterns. The model reproduces the timing differences observed when inhibition or excitation is selectively removed. Image adapted from Fischl et al. (2024).
Gene Therapy Restores Mechanical Fidelity for Timing Input
With timing and over-suppression handled, how do we maintain that the input signal is transduced? If the brainstem refines timing, the cochlea creates it – it’s here where hair cells transduce mechanical deflections into graded receptor potentials. However, when its mechanics break down, precision fails everywhere. chloride intracellular channel 5 (CLIC5) is a gene linked to recessive deafness and vestibular dysfunction in humans. In Clic5-deficient mice, the stereocilia bundles become disorganized and inner ear function collapses. To further explore this finding, the researchers delivered wild-type Clic5 into neonatal mice using adeno-associated virus (AAV) vectors [single-stranded (ssAAV) and self-complementary (scAAV)] (3).
Both constructs restored CLIC5 protein expression at the base of the stereocilia. Treated mice showed preservation of hair bundle morphology and full recovery of auditory brainstem response (ABR) thresholds and vestibular behavior (circling, swimming reflexes). scAAV vectors achieved comparable recovery at lower titers than ssAAV vectors, suggesting improved transduction efficiency and reduced toxicity.
Immunolabeling using our Anti-CLIC5 Antibody (#ACL-025) confirmed correct CLIC5 localization to the stereocilia base and scanning electron microscopy (SEM) imaging showed preserved bundle architecture (Figure 3). Without structural anchoring by CLIC5, force transmission to mechanotransduction channels is degraded, disrupting receptor potential timing and downstream spiking.
The cochlea can only encode time if the hair bundle transmits force fast and cleanly. This therapy restores that structural fidelity. By stabilizing the very first step in sound transduction, the work here shows that timing can be repaired – not just measured. In addition, by achieving full recovery with lower dose vectors, it points toward safer, more feasible gene therapies for inherited hearing and balance disorders.
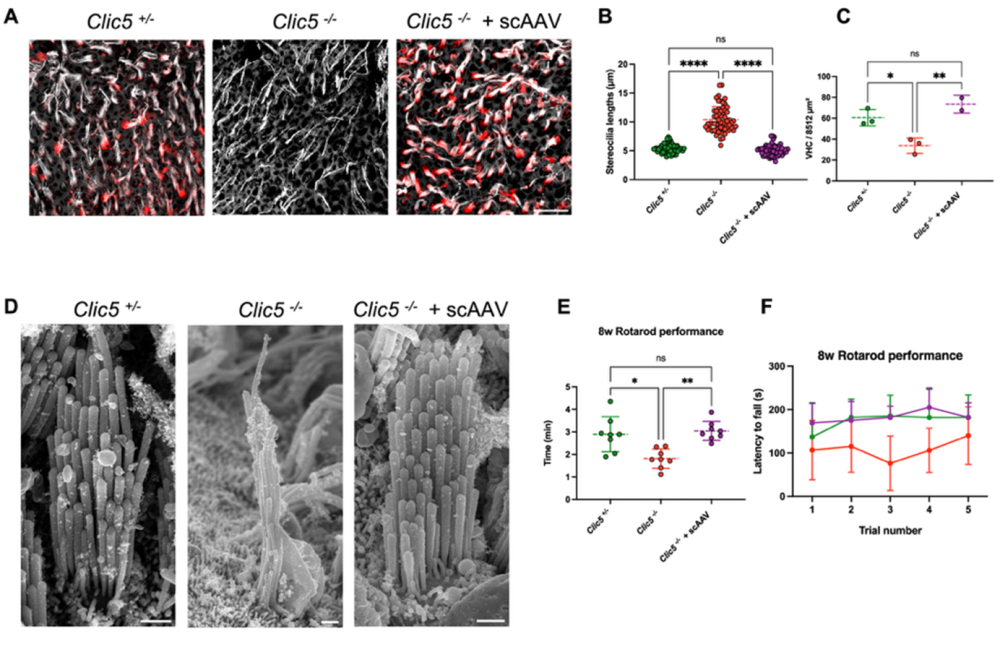
Figure 3. Gene therapy using an adeno-associated virus (AAV) vector restores balance and hair cell structure in chloride intracellular channel 5 (Clic5)-deficient mice. This figure shows how delivering the Clic5 gene with a self-complementary adeno-associated virus (scAAV) can correct defects caused by loss of CLIC5. The CLIC5 is a protein essential for the normal structure and function of vestibular hair cells (VHCs), the sensory cells responsible for balance. A) Confocal microscopy images of VHC in the utricle from three groups of mice: normal (Clic5+/-), Clic5-deficient (Clic5-/-), and Clic5-deficient mice treated with scAAV. Cells were stained for CLIC5 (red) and actin (gray) to visualize the restoration of protein expression and hair bundle morphology. B) Measurements of stereocilia length show that AAV-treated Clic5-/- mice have hair bundles closer to normal length compared to untreated knockouts. C) Quantification of hair cell survival at 12 weeks indicates that scAAV treatment improves cell maintenance in the utricle. D) Representative scanning electron microscopy images of VHCs from the utricle of Clic5+/−, Clic5−/−, and Clic5−/− mice injected with scAAV at 12 weeks. The scale bar represents 10 μm for A) and 1 μm for D). E) Average time spent on the rotarod for Clic5+/-, Clic5-/-, and Clic5-/- mice injected with scAAV (n = 8), assessed at 8 weeks. F) Latency to fall during 5 trials for the same groups shown in E. (D–F) show that viral treatment restores the normal morphology and size of hair bundles. Statistical test for A), B) and E) was one-way ANOVA followed by Tukey correction for multiple comparisons, and for F) – two-way ANOVA with Holm–Sidak correction for multiple comparisons. P values: *P < 0.05, **P < 0.01, ****P < 0.0001, ns= not significant. Image adapted from Hahn et al. (2025).
Timing Loss Activates a Latent Death Pathway in the Auditory Brainstem
But what happens when things go wrong – how does the hearing system respond when there’s a complete loss of input? Researchers from Japan removed afferent input by unilateral cochlear ablation in post-hatch chicks and examined cochlear nucleus neurons over 24 hours (4).
RNA sequencing revealed rapid upregulation of pro-apoptotic p75NTR–JNK pathway genes. Western blotting confirmed persistently high levels of pro brain-derived neurotrophic factor (proBDNF), the unprocessed precursor of BDNF, which also triggered the pro-apoptotic p75NTR–JNK pathway. Within six hours, FLAG-tagged p75NTR had accumulated at the neuronal surface, leading to caspase activation and structural degeneration.
To test causality, the team exposed embryonic brainstem slice cultures to Alomone’s Recombinant human proBDNF (cleavage resistant) protein (#B-256). This resulted in dose-dependent cell death, saturating near 1 ng/mL – a concentration consistent with full p75NTR activation. Blocking synaptic transmission with TTX, CNQX, or D-AP5 (#D-145) – a glutamate receptor antagonists, reproduced this effect, increasing surface p75NTR expression and propidium iodide (PI) labeling. In addition, a dominant-negative p75NTR construct rescued viability. Together, these findings reveal that afferent activity suppresses a latent death pathway. When auditory timing input disappears, intracellular calcium levels decrease, proBDNF–p75NTR signaling dominates, and neurons interpret silence itself as a cue to die (Figure 4).
In the context of hearing as a timing system, this is the endpoint of failure: without ongoing patterned input, the circuit dismantles itself. The same timing precision that encodes sound in the healthy brainstem becomes a dependency – one that must be maintained, electrically or therapeutically, to preserve auditory neurons through deafness.
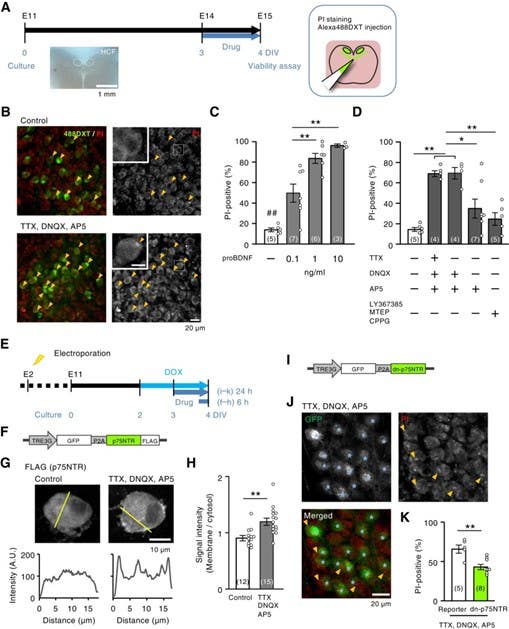
Figure 4. p75NTR signaling mediates neuronal death after activity deprivation. A) Time course of the experiments. Slice cultures were prepared at Embryonic Day 11 (E11), incubated with drugs at day 3 in vitro (3 DIV) for 1 day, and the cell death rate was evaluated at 4 DIV. HCF= Hair Cell Field. B) Cell death assay of nucleus magnocellularis (NM) neurons, the avian equivalent of the mammalian cochlear nucleus. Upper panel: no blockades, lower panel: synaptic and spike activities were blocked by glutamate receptor antagonists: TTX, DNQX, AP5. Dead cells were labeled by propidium Iodide (PI, on the left – red; on the right – gray) before fixation, while NM neurons were visualized by dextran Alexa488 (488DXT, left – green). Squares show magnified areas; arrowheads point to PI-positive cells marking dead neurons. C, D) Effects of proBDNF (C) and blockers of glutamate receptors and sodium channels (D) on the percentage of PI-positive cells. Note that the percentage of PI-positive cells was the highest for antagonists of ionotropic glutamate receptors (DNQX, AP5) but modest for those of metabotropic receptors (LY367385, MTEP, CPPG). E) Time course of the experiments. Plasmids were delivered at E2 using in ovo electroporation, and gene expression was induced at 2.5 DIV by adding doxycycline (DOX, 2 µM), a chemical triggering induction of gene expression. The drugs were applied at 3 DIV. F) Schematic of the DOX-inducible construct used for p75NTR expression. The TRE3G promoter drives co-expression of GFP and FLAG-tagged p75NTR, separated by a self-cleaving P2A sequence, allowing simultaneous visualization (GFP) and detection (FLAG) of the expressed receptor. G) Immunostaining of FLAG-tagged p75NTR in control and activity-blocked neurons. In control cells, p75NTR labeling is distributed throughout the cytoplasm, whereas treatment with TTX, DNQX, and AP5 increases membrane-associated p75NTR. Line-scan intensity profiles (below, yellow lines) show higher fluorescence at the membrane after activity blockade, indicating receptor accumulation at the cell surface. H) Relative signal intensity between membranous and cytoplasmic regions. I) Schematic of the DOX-inducible construct expressing a dominant-negative form of p75NTR (dn-p75NTR). The TRE3G promoter drives co-expression of GFP and dn-p75NTR separated by a P2A peptide, enabling visualization of transfected cells (GFP) and selective inhibition of p75NTR signaling. J) Slice cultures treated with TTX, DNQX, and AP5. Images reveal PI-positive neurons (red, arrowheads) and GFP-positive dn-p75NTR–expressing cells (green). The stars denote reporter-only control cells. K) Quantification of PI-positive neurons demonstrates a significant decrease in cell death in dn-p75NTR–expressing cells relative to reporter controls (P < 0.01). Numbers in parenthesis are the number of cells in (H) and the number of animals in (C, D, K). * P < 0.05, ** P < 0.01 by one-way ANOVA (C, D) or Student’s t-test (H, K). ## represents significant differences between all other groups and the control (p < 0.05) (C). Image adapted from Sato et al. 2025.
A Timeline of Hearing
KV4 channels align spikes to milliseconds. Brainstem inhibition adds strategic delay. Gene therapy restores structure to recover synchrony. Activity itself is required to keep auditory neurons alive.
So what? Three implications that follow from the data:
- Test for timing. Good hearing is more than about simply detecting sound – it’s about when sound arrives. Clinical metrics should reflect that.
- Protect the circuit. Structure matters. So does activity. Preserve both, early and often.
- Build for precision. From gene therapy to hearing tech, success depends on respecting time, and the biology that keeps it.
Hearing is a timing problem solved at multiple scales: molecules (KV4, CLIC5, and proBDNF/p75NTR), cells (bushy and MOC neurons), and circuits (afferent precision vs. efferent control). Lose the beat and you lose the signal; restore the beat and function returns.
Read the first part on smell here and follow us as the 5 Senses series moves from hearing to taste, vision, and touch – decoding how the brain builds perception, one sense at a time.




 protein")
