
A fascinating study has shed light on the dynamic response of vasopressin neurons to osmotic stimuli, focusing on the translocation of ΔN-TRPV1 channels. These channels, when triggered by osmotic changes, move to the surface of vasopressin-releasing neurons. This translocation plays a pivotal role in the neurons’ ability to adapt and respond to varying osmotic conditions, a fundamental process for maintaining fluid balance in the body.
Maintaining Balance
Our bodies constantly regulate the balance of water and salts, a process known as osmoregulation. Central to this process are specialized brain cells – magnocellular neurosecretory cells (MNCs). These cells are responsible for releasing vasopressin, a hormone that plays a crucial role in maintaining our body’s fluid balance by adjusting urine production in response to plasma osmolality changes.
These MNCs, inherently sensitive to osmolality, rely on the mechanical activation of an N-terminal variant of the transient receptor potential vanilloid1 cation channel (ΔN-TRPV1) VP release. Influenced by the cell’s cytoskeleton, these channels respond to osmotic shifts, triggering membrane depolarization and sustained VP secretion.
During sustained osmotic changes, the MNC plasma membrane expands, potentially affecting ΔN-TRPV1 channel activity and neuron excitability. This response may involve other processes, as evidenced by TRPV1 channel translocation in other neurons enhancing excitability, a reaction initiated by phospholipase C (PLC) and protein kinase C (PKC) activation. In MNCs, PLC activation by hypertonic solutions has been shown to be crucial for both MNC hypertrophy and ΔN-TRPV1 activation. This led the researchers to hypothesize that ΔN-TRPV1 trafficking is integral to MNC adaptation under prolonged osmotic stress.
Osmotic stimuli cause ΔN-TRPV1 channels to move to the surface of vasopressin neurons
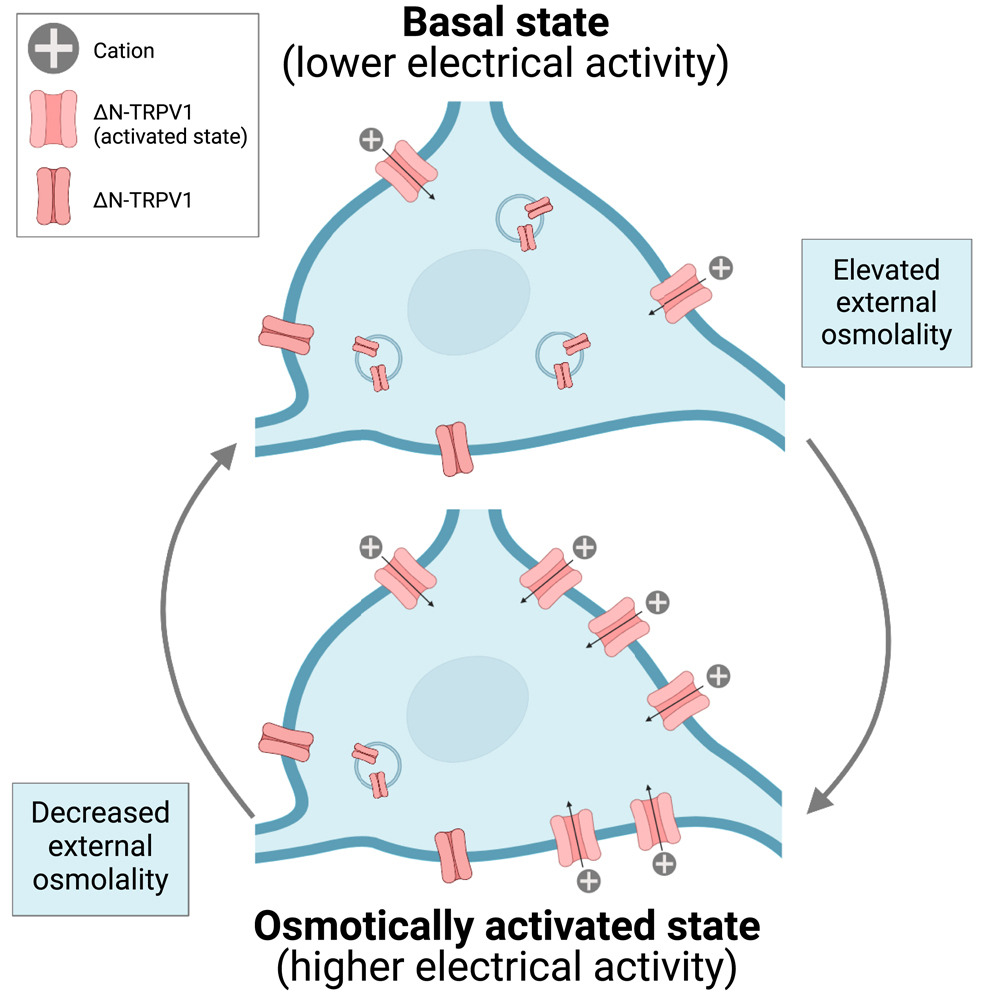
Figure 1. Graphical summary, supporting the idea that Osmotic stimuli cause ΔN-TRPV1 channels to move to the surface of vasopressin neurons. This process is reversible and depends on exocytosis and activation of PLC and PKC. Image from Haan KD et al. iScience. 2023 Feb 20;26(3):106258.
The Remarkable Reversibility of Channel Translocation
What their investigations revealed was that the movement of ΔN-TRPV1 channels in response to osmotic changes isn’t just a transient or one-dimensional response – it’s a reversible process, showing a remarkable level of cellular adaptability. This reversibility is significant because it indicates that vasopressin neurons can modulate their sensitivity to osmotic stimuli dynamically.
When the osmotic pressure returns to normal, the ΔN-TRPV1 channels retreat back into the cell, ready to be deployed again when needed. This back-and-forth movement is a fine example of how cellular components can be recycled and reused in response to environmental changes, ensuring efficient and effective cellular functioning.
The Intricate Process of Exocytosis in Channel Translocation
Exocytosis plays a central role in this translocation process. Typically understood as a way cells expel materials, in this context, exocytosis serves a different purpose: it drives the movement of ΔN-TRPV1 channels to the cell surface in response to osmotic stress. This process involves the fusion of vesicles containing these channels with the plasma membrane, effectively embedding the channels in the membrane where they can function. The regulated nature of this exocytosis process is key to the precision with which neurons respond to osmotic changes, which ensures the right amount of channels are transported to the surface at the right time.
Enzymatic Control: The Roles of Phospholipase and Protein kinase enzymes
The activation of specific enzymes, PLC and PKC, is critical to this process. These enzymes aren’t just simple catalysts; they act as key regulators in the translocation of ΔN-TRPV1 channels. PLC, upon activation, leads to the production of diacylglycerol (DAG) and inositol trisphosphate (IP3), which in turn activate PKC. PKC then aid the process of exocytosis, effectively linking the enzymatic activity to the physical translocation of the channels. This cascade of enzymatic reactions showcases the complexity and precision of intracellular signaling pathways and their role in managing cellular responses to external stimuli.
The research here also demonstrates the indispensable role of phospholipase C delta 1 (PLCδ1), a central enzyme in the process of neuronal adaptation to osmotic changes. Other than the author’s prior work on MNCs, little was known about the role of PLCδ1 in neurons. In these most recent experiments, using PLCδ1 knockout mice, the researchers observed a notable absence of ΔN-TRPV1 translocation in vasopressin neurons. This finding is particularly significant as it underscores the integral role of PLCδ1 in the process of ΔN-TRPV1 channel translocation.
PLCδ1 is known for its role in catalyzing the breakdown of phosphatidylinositol 4,5-bisphosphate (PIP2) into diacylglycerol (DAG) and inositol trisphosphate (IP3), two molecules that play vital roles in intracellular signaling. The absence of PLCδ1 disrupts this pathway, leading to a failure in the translocation of ΔN-TRPV1 channels to the plasma membrane of vasopressin neurons. This translocation is essential for the neurons to respond effectively to changes in osmotic conditions, particularly in regulating vasopressin release, which is crucial for maintaining fluid balance in the body.
The absence of ΔN-TRPV1 translocation in PLCδ1-deficient mice suggests that PLCδ1 is not merely a participant in this cellular process but a key regulator. Without PLCδ1, the vasopressin neurons cannot adapt their ΔN-TRPV1 channels in response to osmotic stress, which could have significant implications for understanding disorders related to osmoregulation.
ΔN-TRPV1 in Water-Deprivation Conditions
The study delves deeper into the behavior of ΔN-TRPV1 channels under conditions simulating water deprivation, where the authors made continued use of live-cell immunocytochemistry with the Alomone Labs antibody (ACC-029) against the external epitope on the TRPV1 channel (Figure 2). In this scenario, both rats and mice displayed a pronounced increase in the translocation of these channels to the surfaces of their vasopressin neurons. This finding is particularly noteworthy as it highlights how vasopressin neurons, integral to the body’s fluid regulation, amplify their responsiveness under osmotic stress. The increase in channel translocation in these water-deprived animals is a testament to the neuron’s ability to adapt to changes in the body’s hydration state.
MNCs deprived of water for 24 h showed a large and reversible increase in size and ΔN-TRPV1 immunofluorescence
Figure 2. (A) Representative images showing TRPV1 immunofluorescence in MNCs isolated from water-deprived rats in the listed conditions. (B) Bar-scatter plots showing the cross-sectional area (CSA; μm2; left) and TRPV1 immunofluorescence labeling in arbitrary units (a.u.) in the MNC plasma membrane (right) in the listed conditions. (C) Bar-scatter plots showing the changes in density of TRPV immunofluorescence on the MNC plasma membrane (a.u. per μm; left) and the change in internal TRPV1 immunofluorescence (a.u.; right) in the listed conditions. Image and legend from Haan KD et al. iScience. 2023 Feb 20;26(3):106258.
The observation of enhanced ΔN-TRPV1 translocation in water-deprived rats and mice is a clear indicator that vasopressin neurons have a built-in mechanism to heighten their sensitivity under dehydration conditions. This heightened sensitivity ensures that the body can respond more efficiently to osmotic imbalances that occur during dehydration. The translocation of these channels is not just a passive response but a proactive adaptation, allowing the neurons to be more effective in managing osmoregulatory challenges.
The increased movement of ΔN-TRPV1 channels to the neuron surface in conditions of water deprivation suggests a potential feedback mechanism. This mechanism could be vital for the regulation of vasopressin release, thereby playing a major role in how the body manages fluid balance under stress. It also underscores the plasticity of vasopressin neurons, able to modify their functional properties in response to the body’s hydration status.
Bridging the Gap to Clinical Relevance
Understanding this adaptive mechanism in vasopressin neurons has clinical implications. In medical conditions characterized by fluid imbalances, such as certain forms of diabetes or conditions involving disrupted thirst perception, these findings could pave the way for new therapeutic strategies. Targeting the regulation of ΔN-TRPV1 translocation might offer novel approaches to modulate vasopressin release, providing a potential avenue for intervention in disorders of water balance.
Concluding Thoughts
This research gives us a new look into the intricacies of how vasopressin neurons, necessary for our body’s fluid balance, adapt to osmotic changes. The reversible translocation of ΔN-TRPV1 channels, contingent on exocytosis and enzyme activation, elegantly demonstrates the dynamic nature of cellular responses. Furthermore, the essential role of PLCδ1 in this process opens new avenues for understanding and potentially manipulating this mechanism in pathological states where fluid balance is disrupted.
Reference
Antibody
Blockers/Antagonists: small molecules
Activators/Agonists: small molecules
Explorer kits
- Extracellular-Targeted TRPV Channel Antibody Explorer Kit (#AK-485)
- pH-Sensitive TRP Channel Antibody Explorer Kit (#AK-475)
