Transient receptor potential vanilloid 1 (TRPV1), a heat- and capsaicin-activated ion channel, drives nociceptor sensitization and chronic pain. Activated by protons, lipids, toxins, or inflammatory mediators, TRPV1 increases calcium influx, reshaping sensory neuron excitability and fueling neuroinflammation. Existing multiplex approaches have often relied on custom-labeled TRPV1 probes or indirect detection. But now, we’ve developed a range of fluorophore-conjugated antibodies and toxins that expand practical, off-the-shelf options.
How TRPV1 Drives and Resolves Pain
TRPV1 acts as a pain switch. Sensitization – by PKC/PKA phosphorylation, extracellular acidification, or inflammatory lipids – promotes calcium influx and nociceptor hyperexcitability. This sensitization drives hyperalgesia in chemotherapy-induced or diabetic neuropathy models, where TRPV1 upregulation is observed in dorsal root ganglia (DRG) (1).
But prolonged activation flips the switch. Capsaicin or resiniferatoxin (RTX) application leads to calcium overload and TRPV1 desensitization, which reduces pain signaling (2, 3). Tau-theraphotoxin-Hs1a (DkTx) targets TRPV1 and locks the channel open, giving you options for functional and structural studies, while RTX has been used experimentally to ablate TRPV1-positive fibers (4).
How does it really work? TRPV1-specific reagents conjugated to a fluorophore in the far-red spectrum to your experiments: the Anti-TRPV1 (VR1)-ATTO Fluor-647N Antibody (#ACC-030-FRN) (Figure 1) and the DkTx-ATTO Fluor-647N (#STD-010-FRN) (Figure 2). The antibody is IHC-, ICC-, and flow-ready in mouse, rat, and human samples, with far-red emission (~670 nm) for low-background imaging. DkTx is a selective toxin that binds TRPV1 specifically, making it useful for studying channel distribution, gating, and desensitization dynamics in live cells (5).
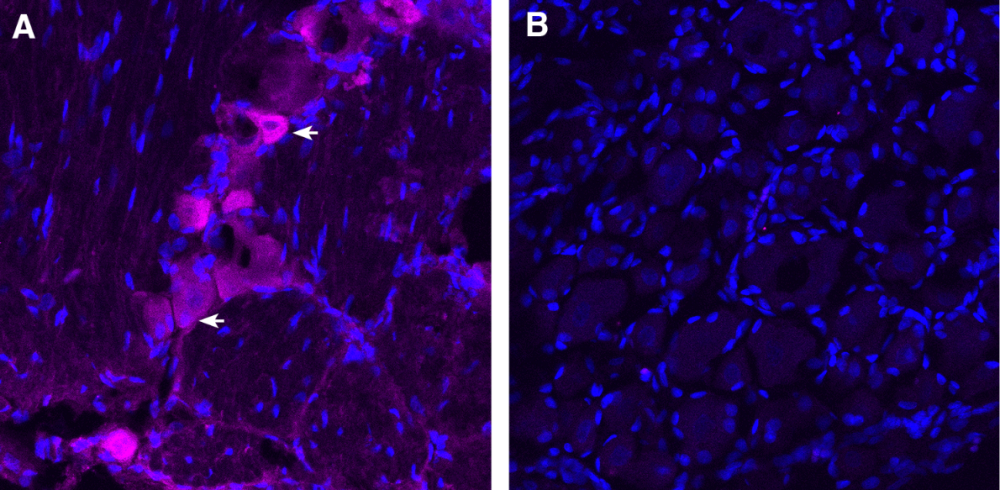
Figure 1. Expression of TRPV1 in dorsal root ganglion (DRG). Frozen DRG sections from a paraformaldehyde-perfused rat were incubated with Anti-TRPV1 (VR1)-ATTO Fluor-647N Antibody (#ACC-030-FRN), (1:100). A) TRPV1 immunoreactivity (magenta) was observed in some DRG cells (arrows). B) Pre-incubation of the antibody with TRPV1/VR1 Blocking Peptide (BLP-CC030), suppressed staining. Cell nuclei are stained with DAPI (blue).
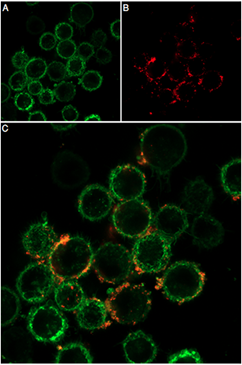
Figure 2. Live cell imaging of DkTx-ATTO Fluor-647N in live intact mouse BV-2 microglia cells. A) CellMask™ Actin 1X solution used to visualize cell membranes. B) The same cells underwent incubation with 0.1 µM of DkTx-ATTO Fluor-647N (#STD-010-FRN), leading to red fluorescence. C) Merged image.
TRPV1 Doesn’t Act Alone in Pain
Multiparameter TRPV1 experiments, with markers like anti-P2Y12 and anti-xCT, allow you to track how neuroimmune crosstalk and oxidative stress shape nociceptor behavior in chronic pain.
That’s why we’ve also developed several mFluor™ Violet 450-conjugated antibodies that target additional pain-relevant systems:
- Anti-P2Y12 Receptor (extracellular)-mFluor™ Violet 450 Antibody (#APR-020-V): P2Y12R is a microglial GPCR that regulates neuropathic pain via p38 MAPK and RhoA/ROCK2 pathways (6, 7).
- Anti-β1-Adrenergic Receptor (extracellular)-mFluor™ Violet 450 Antibody (#AAR-023-V): β1-adrenergic receptor is a sympathetic effector that likely plays a role in experimental hyperalgesia and clinical pain in certain cases (8), although its role is primarily in cardiac function (9).
- Anti-xCT/SLC7A11 (extracellular)-mFluor™ Violet 450 Antibody (#ANT-111-V): xCT/SLC7A11 is glutamate/cystine antiporter controlling oxidative stress and redox balance, which are implicated in excitotoxicity and oxidative stress in chronic and neuropathic pain conditions (10).
Multiplex panels that combine far-red TRPV1 tools with violet-labeled neuroimmune and metabolic markers give you more opportunities to map cellular crosstalk in pain.
Application Highlights
Using DRG neurons or spinal cord microglia from neuropathic pain models, you can track TRPV1 expression and functional states with ATTO Fluor-647N conjugated antibodies and toxins. Simultaneously, mFluor™ Violet 450-conjugated antibodies targeting P2Y12, β1-adrenergic receptor, and xCT allow detailed phenotyping of microglial activation, sympathetic modulation, and glutamate/redox homeostasis within pain circuits.
Multiplex approaches like this let you look deeper into
- TRPV1’s modulation across neuronal and non-neuronal compartments
- Microglia-mediated neuroinflammation underlying pain hypersensitivity
- Sympathetic–immune interactions contributing to chronic pain
- Excitotoxic and oxidative stress mechanisms in pain pathogenesis
Multiplexing cuts background by eliminating secondary antibodies (read all about it) and supports high-content phenotyping: immune–neuronal interaction, oxidative stress, and TRPV1 modulation, all in one experiment.
Limitations and Prospects
While violet-conjugated TRPV1 antibodies haven’t been developed yet, we do offer custom conjugation, making panel expansion feasible. Early tests show excellent performance of DkTx probes across DRG cells, microglia, and heterologous systems, supporting their broad applicability.
Spectrally distinct, validated antibodies allow for precise dissection of pain circuits, linking TRPV1 dynamics, neuroimmune signaling, and redox stress in chronic pain models. And if you need a technical question answered, or a custom reagent developed, we’re always on hand to help.

-ATTO Fluor-647N Antibody")

