Orphan G protein-coupled receptors (GPCRs) are a subset of the GPCR superfamily with no identified endogenous ligands. Only recently, limited functional characterization has been reported. Many are highly conserved and expressed in central nervous system (CNS) regions affected by neurodegenerative disease. Several class A orphan GPCRs previously dismissed as ‘undruggable’ are now being investigated as disease-modifying targets in Alzheimer’s disease (AD), Huntington’s disease (HD), Parkinson’s disease (PD), and multiple sclerosis (MS). New structural tools, ligand screens, and pathology models are changing how we view these receptors (1, 2). The most promising candidates include GPR17, GPR37, GPR52, and GPR88 GPCRs – each with emerging mechanistic roles in synaptic regulation, glial activation, or protein aggregation.
Until recently, orphan GPCRs were largely ignored due to the absence of endogenous ligands and lack of clear pathway readouts. However, two discoveries overturned this view: cryo-electron microscopy (cryo-EM) and structural modelling (3) have allowed virtual screening and structure-based drug design even without a known ligand. Moreover, neurodegenerative phenotypes in GPCR knockout models (4) have revealed non-redundant roles for specific orphan GPCRs in glial and neuronal survival.
These data resulted in a shift from descriptive transcriptomics toward mechanistic investigation and therapeutic targeting. Here, we’ll review some of these receptors, along with our available antibodies, which can be used to study them.
GPR17: Oligodendrocyte Brake in MS and Beyond
GPR17 is transiently expressed in oligodendrocyte precursor cells (OPCs), where it acts as a negative regulator of maturation (4, 5). In mouse experimental autoimmune encephalomyelitis (EAE) models and MS lesions, GPR17 upregulation correlates with stalled differentiation. In contrast, genetic deletion or pharmacological inhibition of this GPCR, promotes myelin repair. From a therapeutic perspective, antagonists could remove this ‘brake’ on remyelination, with particular relevance in relapsing-remitting MS. However, deletion or pharmacological inhibition alone only seem to have modest effects and may not robustly enhance repair in all models (6).
Reagent
- Anti-GPR17 Antibody (#AGR-048) (Figure 1)
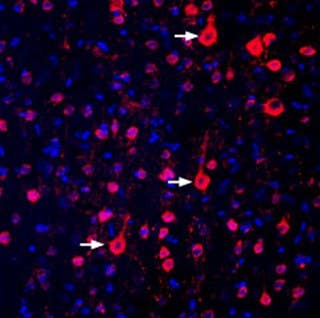
Figure 1. Expression of GPR17 in the rat parietal cortex. Immunohistochemical staining of perfusion-fixed frozen rat brain sections with Anti-GPR17 Antibody (#AGR-048), (1:300), followed by donkey anti-rabbit-biotin antibody and streptavidin-Cy3. GPR17 immunoreactivity (red) appears in the cortical cells (arrows). The cell nuclei were stained with DAPI (blue).
GPR37: PD Relevance, via Glial Stress Buffering
GPR37 is highly expressed in glial cells and dopaminergic neurons. In PD models, it interacts with parkin, modulates ER stress responses, and influences dopaminergic survival. Knockout mice show increased sensitivity to MPTP neurotoxicity, which suggests a protective role (7). Despite being an orphan, GPR37 traffics abnormally in PD and is found aggregated in Lewy bodies, raising questions about its role in protein homeostasis.
Reagents
- Anti-GPR37 (extracellular) Antibody (#AGR-049)
- Anti-GPR37L1 (extracellular) Antibody (#AGR-050) (Figure 2)
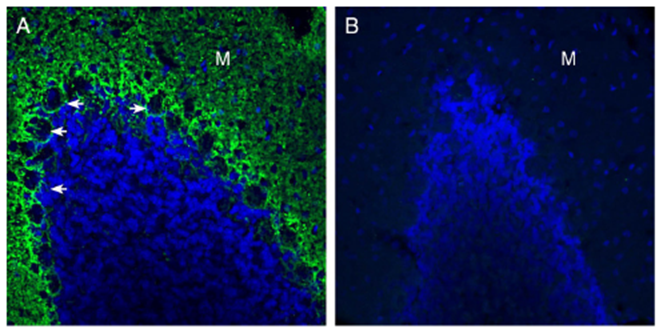
Figure 2. Expression of GPR37-Like 1 (GPR37L1) in the rat cerebellum. Immunohistochemical staining of perfusion-fixed frozen rat brain sections with Anti-GPR37L1 (extracellular) Antibody (#AGR-050), (1:300), followed by goat anti-rabbit-AlexaFluor-488. A) GPR37L1 immunoreactivity (green) appeared around Purkinje cell soma (arrows) and in the molecular layer (M). B) Pre-incubation of the antibody with GPR37L1 (extracellular) Blocking Peptide (#BLP-GR050), suppressed the staining. The cell nuclei were stained with DAPI (blue).
GPR52 and GPR88: Modulating Dopamine in HD and Schizophrenia
GPR88 is enriched in the striatum and regulates dopamine D2 receptor signaling (8) . Knockout mice show hyperdopaminergic phenotypes, with behavioral traits reminiscent of HD and schizophrenia, although these effects may not be strictly neurochemical (9). In contrast, GPR52 promotes dopamine D1 receptor activity and reduces mutant huntingtin aggregation when activated (10). These receptors are being targeted in early-stage drug programs for HD symptom modulation and behavioral stabilization (2).
Reagents
- Anti-GPR52 Antibody (#AGR-058) (Figure 3)
- Anti-GPR88 Antibody (#AGR-038) (Figure 4)
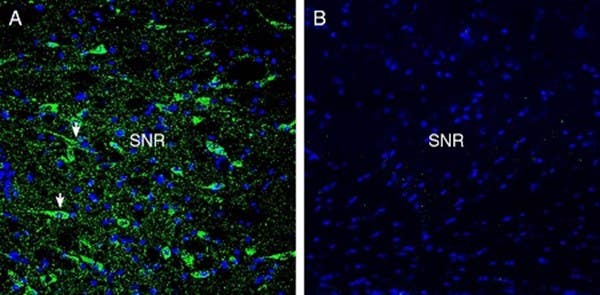
Figure 3. Expression of GPR52 in the rat substantia nigra. Immunohistochemical staining of perfusion-fixed frozen rat brain sections with Anti-GPR52 Antibody (#AGR-058), (1:300), followed by goat anti-rabbit-AlexaFluor-488. A) GPR52 immunoreactivity (green) appeared in the neuronal profiles (arrows). B) Pre-incubation of the antibody with GPR52 Blocking Peptide (#BLP-GR058), suppressed the staining. The cell nuclei were stained with DAPI (blue). SNR = substantia nigra pars reticulata.
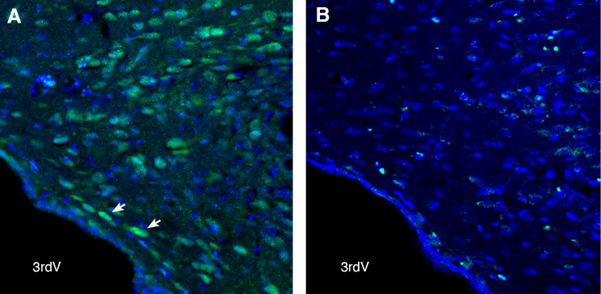
Figure 4. GPR88 in the rat dorsomedial hypothalamus. Free-floating frozen brain sections from a paraformaldehyde-perfused rat were incubated with Anti-GPR88 Antibody (#AGR-038), (1:300), followed by goat anti-rabbit-Alexa Fluor-488 (green). A) GPR88 immunoreactivity was observed in neurons of the dorsomedial hypothalamus (arrows). B) Pre-incubation of the antibody with the GPR88 Blocking Peptide (#BLP-GR038) suppressed the staining. The cell nuclei were stained with DAPI (blue). 3rdV = third ventricle.
Other Neurodegenerative Diseases and Processes
There are more uncharacterized orphan GPCRs out there with potential roles in neurodegenerative diseases and processes (1, 2). Here’s a short list of the orphan GPCRs and the antibodies you can use to target them.
AD: AD-linked orphans primarily modulate neuroinflammation (GPR18 and GPR55), circadian/metabolic context (GPR50), and neuronal survival and other functional outcomes in models of GPR52 and GPR139.
Reagents
Multiple sclerosis and myelin biology: GPR17 links extracellular nucleotides/leukotrienes to oligodendrocyte precursor cell (OPC) state transitions; GPR55 influences inflammatory tone.
Reagents
- Anti-GPR55 (extracellular) Antibody (#ACR-062)
- Anti-GPR55 (extracellular)-FITC Antibody (#ACR-062-F)
Neuroinflammation and glial activation: GPR84 promotes microglial cytokine release, chemotaxis, and phagocytosis. It is upregulated in microglia of the APP/PS1 AD mouse model and differentially expressed in the human AD entorhinal cortex. GPR18 functionally interacts with cannabinoid CB2 receptor, in microglia, and engages pro-resolving signalling in brain injury models. GPR83 is expressed in regulatory T cells and CNS regions and is linked to immune regulation and stress-associated physiology.
Reagents
- Anti-GPR84 (extracellular) Antibody (#AGR-052)
- Anti-GPR84 (extracellular)-APC Antibody (#AGR-052-APC)
- Anti-GPR84 (extracellular)-FITC Antibody (#AGR-052-F)
- Anti-GPR84 (extracellular)-PE Antibody (#AGR-052-PE)
- Anti-GPR18 (extracellular) Antibody (#AGR-068)
What’s Missing: Ligands, Bias, and Context
The key limitation of researching orphan GPCRs remains ligand identification. In some cases, surrogate ligands (e.g. for GPR52) or compounds have been developed, but most class A orphans still lack validated small molecules. This constrains both functional studies and drug development. However, new screening techniques – such as multiplexed antibody selectivity assays, and structural models from cryo-EM or AlphaFold – are closing that gap.
A New Relevance for Disease
These receptors don’t fit the old model of GPCR drug discovery. Most show tissue-restricted expression, interact with disease-relevant pathways, and modulate glial–neuronal crosstalk – an emerging therapeutic interaction in CNS disease. They may not function as classical synaptic transmission modulators but may rather contribute to the regulation of stress responses, cell survival, and inflammatory thresholds. As antibodies and chemical probes become more available, these orphans are likely to move from transcriptomic curiosities to mechanistic nodes. Their relevance to disease moves from speculative to increasingly measurable.

 Antibody")
 Antibody")
 Blocking Peptide")





 Antibody")
 Antibody")
 Antibody")