How fluorescently conjugated α-bungarotoxin (BTX) is being used as a precise neuromuscular junction (NMJ) marker, without blocking synaptic function.
BTX is usually utilized as a pharmacological tool. It is a snake venom-derived peptide that binds nicotinic acetylcholine receptors (nAChRs) with near-irreversible affinity and blocks neuromuscular transmission. However, in a growing number of imaging studies, BTX is being used in a very different way – one that has little to do with blocking and everything to do with marking.
In many neuromuscular junction (NMJ) imaging studies, fluorescently conjugated α-bungarotoxin (BTX) is used exclusively as a structural marker in fixed tissue preparations. In this context, synaptic transmission is no longer occurring, and BTX is not applied during live or functional assays. Instead of acting as a pharmacological blocker, it serves as a highly specific labeling reagent that defines the postsynaptic acetylcholine receptor clusters with precision. This distinction is essential for understanding its widespread use in studies focused on morphology, localization, and synaptic organization rather than receptor inhibition.
Defining the NMJ as a Region of Interest
At the NMJ, postsynaptic nAChRs are densely clustered in a highly stereotyped pattern. BTX binds these receptors with exceptional specificity and affinity, to give a clean, high-contrast outline of the postsynaptic endplate. Once BTX is conjugated to a fluorophore, it becomes an unusually reliable way to visualize the NMJ, particularly in muscle tissue where background staining can otherwise be a challenge. Conjugation to bright, photostable fluorophores further enhances its value, enabling high-contrast visualization of acetylcholine receptor clusters even in complex muscle tissue.
In this context, BTX functions as a positional reference. It tells the experimenter where the NMJ is, so that other signals such as those representing presynaptic proteins, axonal pathology, local translation, or synaptic alignment can be measured relative to that location. When used in fixed samples, fluorescent α-bungarotoxin is not a pharmacological intervention but a structural tool. Its high-affinity binding to nicotinic acetylcholine receptors makes it uniquely suited to define the postsynaptic architecture of the NMJ with clarity and consistency, without influencing the biological processes under investigation. By separating functional experiments from post-fixation labeling, researchers ensure that receptor blockade plays no role in their readouts. In this imaging context, BTX is best understood not as a toxin acting on synaptic physiology, but as a precise molecular tool that enables accurate spatial analysis of neuromuscular structure.
Figure 1 shows that in amyotrophic lateral sclerosis (ALS) models, α-Bungarotoxin-ATTO Fluor-633 (#B-100-FR) is used to delineate NMJs. This allows for the quantification of the presynaptic accumulation of TAR DNA-binding protein 43 (TDP-43) and changes in local protein synthesis within a precisely defined synaptic region (1).
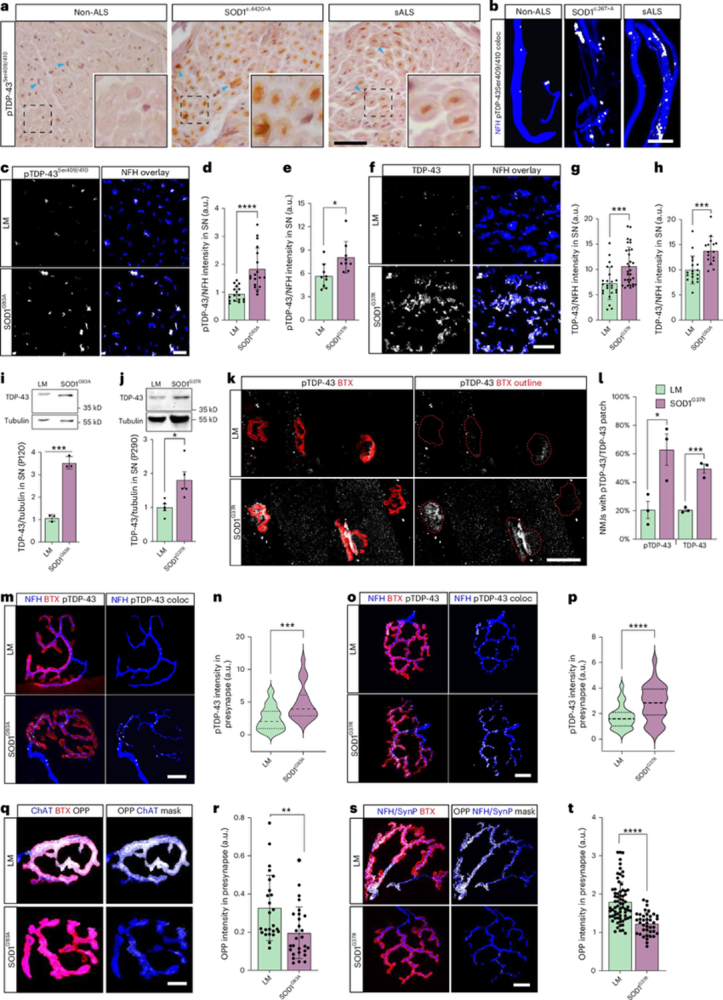
Figure 1. TDP-43 peripheral pathology in SOD1 patients and SOD1 mouse models. (a) Immunohistochemical staining of pTDP-43 in obturator nerve biopsies of non-ALS patients (non-ALS; motor neuropathy), patients with SOD1 (c.442 G > A mutation) and patients with sporadic ALS (sALS). Arrowheads indicate Schwann cells. (b) Immunofluorescent staining of pTDP-43 in intramuscular nerves in biopsies of non-ALS patients, patients with SOD1 (c.26 T > A mutation) and patients with sALS. White indicates Neurofilament Heavy Chain (NFH)–pTDP-43 co-localization (coloc). (c–e), Immunofluorescent images and quantification of pTDP-43 in sciatic nerve (SN) cross-sections of SOD1G93A (d) and SOD1G37R (e) mice and their littermates (LMs). White indicates NFH-masked TDP-43. (d). (f–h) Immunofluorescent images and quantification of TDP-43 in SN cross-sections of SOD1G37R (g) and SOD1G93A (h) mice and their LMs. White indicates NFH-masked TDP-43. (i, j), Representative images and quantification of Western blots for TDP-43 (43 kD) and tubulin (55 kD) in SN axoplasms of SOD1G93A (i) and SOD1G37R (j) mice versus their LMs. (k, l), Representative images and quantitative analysis for the percent of NMJs with an apparent TDP-43/pTDP-43 patch in extensor digitorum longus (EDL) muscles from P290 SOD1G37R and LM controls. The red dashed line marks the bungarotoxin (BTX) perimeter. (m–p), Immunofluorescent images and quantification of NFH–pTDP-43 coloc in NMJs of presymptomatic (P60) SOD1G93A (m, n) and (P290) SOD1G37R (o, p) mice versus their LMs. White indicates NFH–pTDP-43 coloc. (q–t), Representative images and quantification of the O-propargyl puromycin (OPP) labeling in NMJs of presymptomatic (P60) SOD1G93A (q, r) and (P290) SOD1G37R (s, t) mice as well as their LMs. White indicates OPP–ChAT or OPP–NFH–synaptophysin three-dimensional coloc.
Image adapted from Ionescu et al. (2025). https://doi.org/10.1038/s41593-025-02062-6. Licensed under Creative Commons Attribution 4.0 International (CC BY 4.0).
Figure 2 shows that in muscular dystrophy models, α-Bungarotoxin-ATTO Fluor-488 (#B-100-AG) outlines the postsynaptic membrane to enable measurements of NMJ morphology and alignment with presynaptic terminals (2).
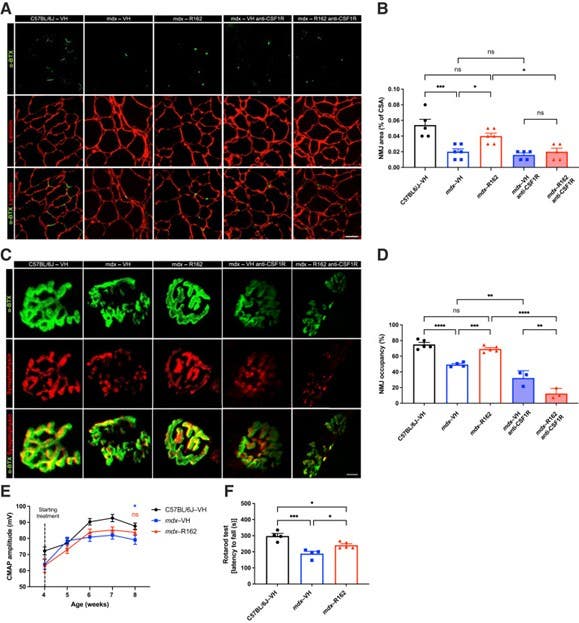
Figure 2. Pharmacologic glutamate dehydrogenase 1 (GLUD1) inhibition restores neuromuscular junction (NMJ) morphology and function in Duchenne muscular dystrophy (mdx) model mice. (A, B), Representative images (A) and quantitative analysis (B) of the NMJ area in cryosections of tibialis anterior muscle from vehicle (VH)-treated C57BL/6J mice and VH- and R162 (an inhibitor of GLUD1)-treated mdx mice in macrophage-competent and -depleted conditions. (C, D), Representative images of NMJs in whole-mount extensor digitorum longus (EDL) muscle showing the presynaptic terminal bouton (synaptophysin) and the postsynaptic acetylcholine receptors [α-bungarotoxin (α-BTX)] (C) as well as NMJ occupancy (D), calculated as (presynaptic area/postsynaptic area) × 100, in EDL muscle from VH-treated C57BL/6J mice and VH- and R162-treated mdx mice in macrophage-competent and -depleted conditions. (E), Compound muscle action potential (CMAP) analysis in VH-treated C57BL/6J mice and VH- and R162-treated mdx mice over 4 weeks of treatment. (F), Rotarod test in C57BL/6J and mdx mice treated with VH or R162 for 4 weeks, followed by 3 weeks of treatment interruption. The data are expressed as the means ± s.e.m. from at least two independent experiments. n = 3 (D, macrophage-depleted mdx–VH and mdx–R162 anti–CSF1R); n = 4 (D, macrophage-competent mdx–VH; F, VH); n = 5 (B, macrophage-depleted mdx–VH and mdx–R162 anti-CSF1R, and C57BL/6J-VH; D, C57BL/6J-VH, and macrophage-competent mdx–R162; F, R162); n = 6 (B, macrophage-competent mdx–VH and mdx–R162); n = 9 (E, C57BL/6J-VH); n = 10 (E, mdx–VH); n = 12 (E, mdx–R162). ∗P < 0.05, ∗∗P < 0.01, ∗∗∗P < 0.001, and ∗∗∗∗P < 0.0001, assessed by one-way ANOVA with the Tukey multiple comparisons test (B, D, and F) and two-way ANOVA with the Bonferroni multiple comparisons test (E). ns, not significant.
Image adapted from Pereira-Nunes et al. (2025). http://doi.org/10.1016/j.ajpath.2025.05.003. Licensed under Creative Commons Attribution 4.0 International (CC BY 4.0).
In both cases, functional readouts – motor performance, muscle contraction, or neurotransmission – are assessed separately, without the toxin being part of the causal chain under investigation.
The fact that BTX is reserved for imaging, after fixation, makes clear that its value here lies in its binding specificity. Functional assays – muscle contraction, grip strength, electrophysiology, or behavioral readouts – are conducted separately and never in the presence of the toxin. This separation ensures that receptor blockade does not influence the biological processes under investigation.
Toxins as Exceptional Markers
In this application of BTX as a marker, conjugated BTX isn’t so different from a directly conjugated antibody. Both are single-step reagents that bind a defined target and generate a spatial signal. The difference is that BTX binds a functional receptor site rather than an antibody epitope, which in some contexts makes it even more reliable. nAChR clusters at the NMJ are abundant, stable, and structurally conserved, and BTX binding is remarkably consistent across preparations.
This reliability is one reason BTX has become something close to a gold standard for NMJ visualization. It labels exactly the structure researchers care about, with minimal ambiguity and without the need for secondary detection or signal amplification. When the goal is to define a region rather than manipulate it, those properties matter more than the molecule’s biological origin.
Of course, BTX does not stop being a toxin simply because it is conjugated to a fluorophore. In live-cell or functional experiments, its ability to block nAChRs remains central, and concentrations, timing, and interpretation must be handled accordingly. But that’s a different experimental mode. In fixed tissue imaging, the same molecule is repurposed and its biological activity becomes largely irrelevant. When the question is spatial rather than functional, BTX moves from blocker to landmark.
And sometimes, the most effective way to see a structure clearly is to take the toxin out of the toxin.