
Vision begins with light – photons striking the retina – but the work of seeing is attributed to membrane proteins. These proteins capture, relay, and renew the molecular signals that turn light into neural code to give us something we can see and understand. From the first flicker of rhodopsin activation to the regulated trafficking of the enzymes that shape phototransduction, membrane proteins create the conditions for perception.
Light starts this process when rhodopsin absorbs a photon in the photoreceptor outer segment – the stacked membrane compartment packed with the machinery for phototransduction. The 11-cis-retinal chromophore isomerizes to all-trans in under a picosecond, and that single structural shift pushes rhodopsin into its active state. Activated rhodopsin then initiates G-protein signaling that lowers cGMP levels, closes cGMP-gated channels, and hyperpolarizes the photoreceptor (1). Membrane proteins then drive recovery: trafficking clears activated components and delivers replacements so the cell can respond to the next photon with the same precision.
In this instalment of our Senses series, we look at vision, specifically how light triggers a tightly choreographed cascade in the eye, and how that process depends on precise protein localization and turnover. Finally, we discuss how receptors could be very interesting new drug targets.
Guiding Water and Glycerol to Keep the Lens Clear
Before light can reach rhodopsin, it has to pass through a lens that remains clear. But it can only achieve this because its membrane channels keep water and solutes exactly where they need to be, thanks to balance maintained by aquaporins (AQPs) – membrane channels that regulate water and solute flow. Among them, AQP3 stands out for transporting not just water but glycerol and hydrogen peroxide to support osmotic stability and redox control.
Recent data confirmed AQP3 expression across mouse, rat, bovine, and human lenses, with two biochemically distinct forms: a non-glycosylated 25 kDa variant enriched in the epithelium, and a 37 kDa glycosylated form found deeper in the inner cortex and core (2). This layered expression suggests zone-specific functions, tuned to the physiological demands of each lens region.
AQP3 localization shifts with age (Figure 1). In mouse embryos and early postnatal stages, it sits in cytoplasmic vesicles. With help from our Anti-Aquaporin 3 Antibody (#AQP-003), researchers observed that by postnatal day 15, AQP3 moves to the plasma membrane of mature fiber cells just as the lens begins to reach its full optical clarity. Rather than just moving water, AQP3 helps the lens grow into a precise, light-focusing structure.
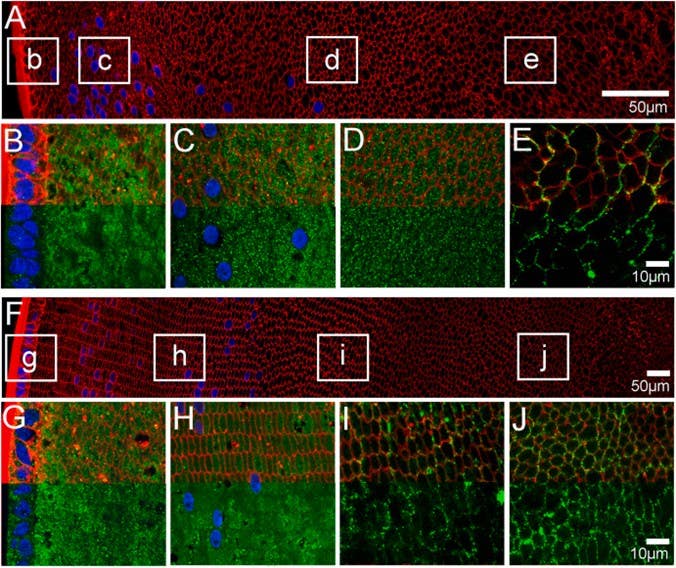
Figure 1. Subcellular localization of aquaporin (AQP)3 at postnatal day 6 (P6) and P15 mouse lenses. A & F) Overview image montage of sections labelled with the membrane marker wheat germ agglutinin (WGA; red) and the nuclei marker DAPI (blue) to show the overall structure of the P6 (A) and P15 (F) mouse lenses sectioned in an axial and equatorial orientation, respectively. The white boxes designate regions in the lens from which high magnification images (B-E & G-J) were obtained to examine the subcellular localization of AQP3 protein (green). B-E) In the epithelium and fiber cells of the peripheral outer cortex of P6 mouse lenses, AQP3 labelling was highly cytoplasmic (B), and this cytoplasmic labelling remained in both the deeper outer cortex (C) and the inner cortex (D). AQP3 expression in the lens core (E) was found to be strongly associated with the membranes (E). G-J) In the epithelium and fiber cells of the peripheral outer cortex of P15 lenses, AQP3 was highly cytoplasmic (G), and this cytoplasmic labelling remained in the deeper outer cortex (H). However, there appears to be less staining, moving from the deeper outer cortex to the inner cortex, where AQP3 is associated with the membranes (I). AQP3 expression remained membranous in the core (J). In the high-power images (B-E & G-J) the red WGA signal was omitted from the bottom half of the images to allow for a better appreciation of the localization of the AQP3 labelling. Image taken from Petrova et al. (2024).
How Photoreceptors Stay Aligned: The Hidden Role of Syntaxin-3 (STX3)
The theme is the same across tissues: optical performance depends on proteins being moved, sorted, and positioned with absolute fidelity. In photoreceptors, this task is attributed in part to STX3, a SNARE protein that directs rhodopsin and peripherin-2 to the disc membranes. When STX3 is deleted, photoreceptors accumulate mislocalized proteins in the inner segment and degenerate rapidly (7). The effect is an immediate miswiring – a breakdown in the spatial logic of the photoreceptor.
This mislocalization of STX3 has emerged as an early marker of disrupted protein trafficking in retinal disease. Normally confined to the inner segment and synapse, STX3 was found to accumulate abnormally in the outer segments (the compartment that houses disc membranes and light-sensing proteins) in animal models of Bardet–Biedl syndrome (BBS) (Figure 3), a ciliopathy that includes early-onset retinal degeneration (8). This mistargeting contributes to early visual dysfunction and progressive photoreceptor degeneration, underscoring the importance of precise membrane protein sorting in maintaining photoreceptor integrity and light responsiveness. Yet even perfect synaptic scaffolds fail if proteins are delivered to the wrong place.

Figure 2. Changes in retinal structure revealed by multiplex immunohistochemistry. This image compares retinal sections from 5-month-old control and Bardet–Biedl syndrome (BBS)1 knockout mice to visualize how protein expression shifts in a BBS model. Anti-Syntaxin 3 Antibody (#ANR-005) staining detected an increase in syntaxin-3 (gray) in the mutant outer segments. In contrast, the levels of opsin, a key photoreceptor protein detected using the 4D2 antibody (green), was reduced in the mutants. The nuclei were labeled with DAPI (cyan), and cellular membranes were highlighted with the Boron–Dipyrromethene (BODIPY) marker (red). The merged images revealed clear differences between control and mutant retinas, illustrating how protein mislocalization contributes to photoreceptor dysfunction. Image taken from Masek et al. (2022).
While trafficking can be a point of failure, it’s also an opportunity for intervention. Proteins that guide metabolic flux, shuttle solutes, or regulate ionic environments create identifiable points for therapy. Transporters on photoreceptors can be used as molecular handles for targeted drug delivery, and ion channels in the visual cortex can determine whether plasticity is open or closed. Two recent studies illustrated how these membrane proteins can be leveraged to deliver drugs directly to vulnerable cells or to reinstate visual processing in adults.
From Photoreceptor Rescue to Plasticity Renewal
Inherited retinal degenerations (IRDs) such as retinitis pigmentosa and Leber’s congenital amaurosis, feature progressive loss of rods followed by cones, driven in many models by toxic elevations of cGMP. The rd1 mouse model mirrors this process, with early rod death leading to secondary cone degeneration. Effective treatments remain limited and improving drug delivery directly to photoreceptors is a major challenge.
In a transporter-guided delivery study, researchers used the high expression of monocarboxylate transporters MCT1 and MCT2 on photoreceptors to develop liposomes conjugated to monocarboxylates (9). Pyruvate-decorated liposomes showed the strongest photoreceptor uptake, outperforming lactate- and cysteine-linked versions. This uptake was reduced when MCT1/2 were inhibited. Using our Anti-MCT1 (SLC16A1) (extracellular) Antibody (#AMT-011) and Anti-MCT2 Antibody (#AMT-012), the authors verified MCT localization on photoreceptor inner segments (Figure 4). When loaded with the cGMP-analog CN04, pyruvate-decorated liposomes reduced photoreceptor cell death in rd1 explants – an effect free CN04 did not exhibit. These results suggested that transporter-guided liposomes could be used to improve drug access in IRDs where photoreceptors remain present but metabolically stressed.
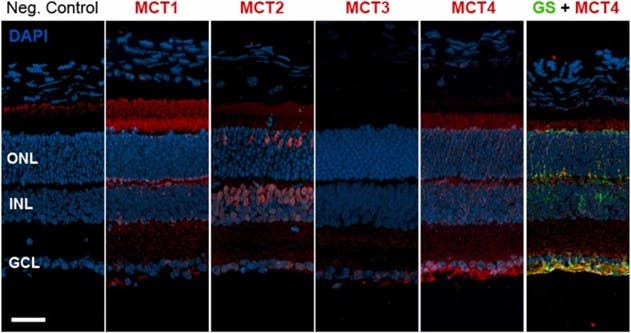
Figure 3. Localization of monocarboxylate transporters (MCTs) in the mouse retina. Retinal cross-sections from wild-type mice (postnatal day 30) were stained to visualize the distribution of four monocarboxylate transporter isoforms: MCT1, MCT2, MCT3, and MCT4 (shown in red). Cell nuclei were labeled with DAPI (blue) to outline the retinal layers, including the ganglion cell layer (GCL), inner nuclear layer (INL), and outer nuclear layer (ONL). To identify Müller glial cells, the sections were also stained for glutamine synthetase (GS; green). MCT4 showed clear colocalization with GS, especially in the Müller cell end-feet, indicating that this transporter is prominently expressed in glial processes. A negative (Neg.) control without the primary antibody is included to show the baseline level of background fluorescence. The scale bar represents 50 µm. Image taken from Christensen et al. (2023).
Another visual system disorder that was examined is amblyopia-often referred to as “lazy eye.” Developing in early childhood, it is the most common cause of vision loss in kids, affecting up to 3 out of every 100 children. Amblyopia arises when abnormal visual experience causes the brain to suppress input from one eye, impairing binocular vision even though the eye itself is structurally normal. In adulthood, recovery is typically limited because cortical plasticity has largely closed. Seeking ways to restore higher-order visual processing when plasticity has diminished, a group of researchers found that increasing brain magnesium using magnesium L-threonate can reinstate multiple measures of plasticity in adult amblyopic mice. These plasticity measures include ocular dominance balance, binocular matching, response reliability, and depth perception (10). Knocking down transient receptor potential melastatin 7 (TRPM7) – a Mg2+-permeable chanzyme – prevented these improvements, as well as the associated increase in NMDA B2 receptor (NR2B) expression. With our Anti-TRPM7 Antibody (#ACC-047), the authors demonstrated that TRPM7 is required for the Mg2+-driven recovery pathway. The findings placed TRPM7 at the center of a mechanism capable of restoring binocular function in adult amblyopia, a condition otherwise resistant to treatment.
Together, these studies show how membrane proteins can be used to both maintain and repair vision – either by targeting degenerating photoreceptors or by re-opening the circuitry required for binocular vision in amblyopia.
Membrane Proteins as Gatekeepers – and Gateways – in Vision
Membrane proteins control every stage of visual signaling: photon capture, water balance, synaptic alignment, cargo trafficking, and now targeted manipulation of cell biology. The molecules reviewed here – rhodopsin, AQP3, STX3, MCT1/2, and TRPM7 – define where signals originate, how they propagate, and whether they can be modified therapeutically. Their localization and turnover determine whether light triggers a clean electrical response or a degraded one, and their accessibility makes them practical targets for intervention.
Reliable reagents remain the foundation for mapping and testing these pathways. Whether identifying transporters on photoreceptors, tracking SNARE mislocalization, or quantifying ion channel-dependent plasticity, validated antibodies and other reagents make it possible to resolve where proteins are, when they move, and how they change in disease or treatment.
This piece continues our Senses series, following earlier posts on taste, hearing, and smell – each driven by membrane proteins turning the physical world into signals. Vision fits the same theme: molecules in motion shaping perception. Next, we close the series with touch, the most intimate sense of all, where force becomes feeling. Stay tuned for Touch: Where Physics Becomes Perception.


 (extracellular) Antibody")

