Extracellular vesicles (EVs) are nanoscale membrane bubbles, ranging from 40 to 1000 nm across, that carry proteins, metabolites, lipids, and RNA between cells. They’re being studied as disease biomarkers, engineered as drug delivery vehicles, and investigated for their role in everything from immune signaling to cardiac repair. But the question of how EVs travel safely has yet to be answered.
They leave one cell, cross the extracellular space, and enter another while moving through ionic environments that shift dramatically. Inside a cell, potassium (K+) sits at around 140 mM. Outside, it drops to roughly 4 mM (a ~30-fold gradient), with a Nernst potential of around −92 mV across the membrane. For a tiny lipid bilayer vesicle carrying a cargo of biologically active molecules, that osmotic swing could be catastrophic. It could swell, burst, and lose the cargo, or shrink and collapse with the same result.
So how do EVs handle it? Databases like ExoCarta have long cataloged ion channels and transporters on EV membranes – specifically, 72 unique ion channels and 107 transporters – but listing proteins in a proteome and showing that they’re functional are very different things. A study published in Nature Communications closed that gap by demonstrating that EVs carry functional, gated, and pharmacologically blockable K+ channels (1).
Catching a Channel in Action
The central technical challenge is obvious: EVs are tiny and standard patch-clamp electrophysiology isn’t compatible with a 200 nm vesicle. The team addressed this in two ways.
First, they used near field electrophysiology (NFE) – a chronoamperometric approach that uses a polypyrrole-based sensor to detect K+ flux from EVs in real time. When EVs were present, K+ currents increased. When iberiotoxin (IBTX) – a highly selective blocker of large-conductance calcium-activated potassium channels (BKCa, encoded by Kcnma1) – was applied, 45% of that current disappeared. From the conductance and open probability assumptions, the authors estimated roughly two functional BKCa channels per vesicle.
That’s electrophysiology conducted in an intact EV, but single-channel properties require a different approach. So, they lysed the EVs, reconstituted the membrane fragments into planar artificial lipid bilayers, and ran classical bilayer recordings. They discovered a K+ channel with a single-channel conductance of 335 ± 15 pS, with ~50% open probability at +40 mV, and a 75% reduction in open probability following IBTX addition at 100 nM. Together, the conductance and pharmacology suggested that they had identified BKCa.
Shifting to in vivo work, the team observed that EVs from Kcnma1⁻/⁻ knockout mice still showed K+ currents in bilayer recordings, but with a much smaller conductance of 110 ± 10 pS – consistent with other K+ channels compensating in the absence of BKCa.
Protein Confirmation of BKCa in EVs
Electrophysiology often reveals a protein’s function, but it is important to know where the protein is localized. Western blotting of plasma-derived EVs from wild-type mice detected BKCa at ~115 kDa. Immunofluorescence, using Alomone’s Anti-KCNMA1 (KCa1.1) Antibody (APC-021), demonstrated high specificity in knockout validation experiments, showing robust labeling of EVs from Kcnma1+/+ mice while yielding no specific signal in EVs derived from Kcnma1⁻/⁻ animals. The antibody showed co-localization with PKH-67-labelled EVs in 72 ± 5% of EVs from Kcnma1+/+ mice, versus no specific signal in EVs from Kcnma1⁻/⁻ animals (Figure 1). The same antibody detected BKCa in 95 ± 2% of EVs purified from mesenchymal stem cell conditioned media. The protein is there, and it’s on the membrane.
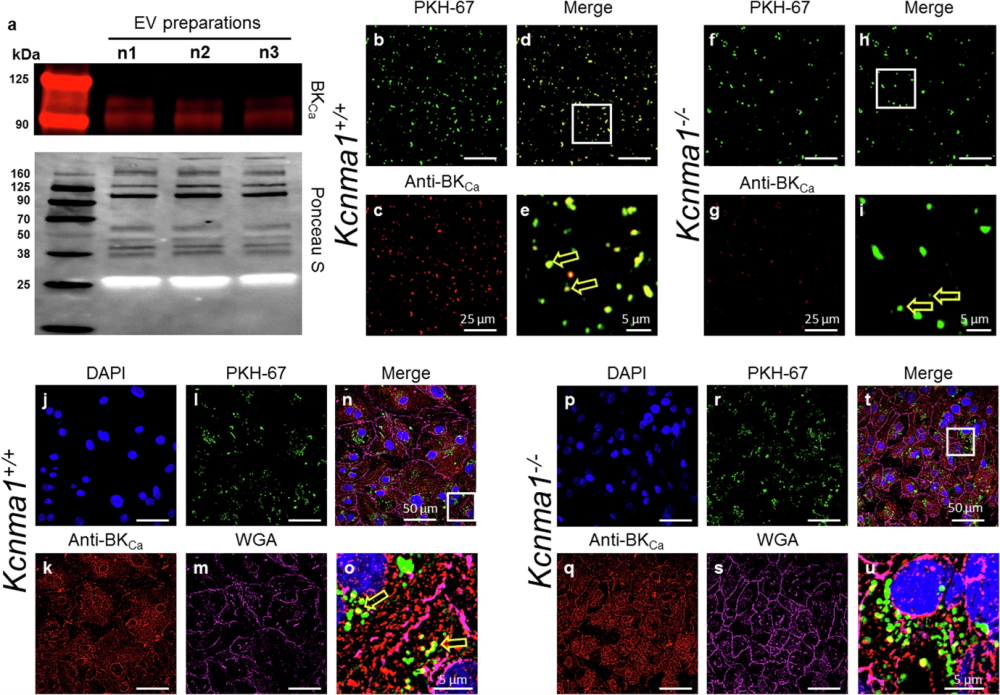
Figure 1. The presence of BKCa in EVs. a Representative Western blot showing the presence of BKCa and the protein loading control (Ponceau S) in EVs isolated from the plasma of mice (n = 3). The migration of molecular weight markers is shown on the left of each panel. b-e Purified EVs from Kcnma1+/+ mice (shown as green, d) and Kcnma1−/− mice (f–i) labeled with PKH-67 (green) or Anti-BKca antibodies (red). e and i are zoomed regions from d and h, respectively, and the arrows in e and i denote presence of BKca channel. All images were merged in d (from b and c) as well as in h (from f and g). Human iPSC-derived cardiomyocytes (hiPSC-CM, j–u) were incubated with EVs isolated from Kcnma1+/+ and Kcnma1−/− labeled PKH-67 (green, l, and r, respectively). hiPSC-CM were loaded with WGA (magenta, m, and s) and fixed for immunocytochemical analysis. hiPSC-CM were permeabilized and labeled with Anti-BKca antibodies (red, k, and q) or DAPI (blue, j and p). All images were merged in n (from j–m) and t (from p–s). o and u are zoomed regions from n and t, respectively. BKCa was present in EVs isolated from Kcnma1+/+ mice (yellow arrow in o) but were not found in Kcnma1−/− mice. b,d,f,h,j-m, and p-s.
Figure taken from Sanghvi et al. (2025). https://doi.org/10.1038/s41467-024-55379-4.
What the BKCa Channel Actually Does
Knowing BKCa is present and functional is a strong foundation, so the next step was to understand ‘why’. Size matters here – literally. The team used nanoparticle tracking analysis (NTA) to measure EV diameter under different K+ conditions. In PBS, wild-type EVs averaged 177 nm. In 4 mM extracellular K+, that dropped to 161 nm. In 145 mM intracellular K+, it rose to 217 nm. The BKCa activator NS1619 pushed the size up slightly, while IBTX pushed it back down (152 nm). EVs from Kcnma1⁻/⁻ mice showed no size differences across K+ concentrations at all. Thus, they lost the ability to respond to the K+ gradient.
There’s a numerical note worth flagging, murine Kcnma1⁻/⁻ plasma contained roughly twice as many EVs as wild-type mice, but they were consistently smaller. The authors interpreted this as a compensatory or dysregulated biogenesis phenotype – though the mechanism isn’t directly established by these experiments.
Cargo Consequences
BKCa status also correlated with what’s inside the vesicle. NanoString miRNA profiling of plasma EVs from wild-type versus Kcnma1⁻/⁻ mice identified 42 differentially expressed miRNAs out of 600 analyzed: 28 were downregulated and 14 were upregulated in knockout mice EVs. Those miRNAs depleted in the absence of BKCa included miR-19a, miR-23a, and miR-486 – miRNAs with established cardioprotective roles. Enriched miRNAs in knockout EVs included miR-15a and miR-144, with rather different downstream effects.
Whether this reflects BKCa directly influencing cargo loading during biogenesis or an indirect consequence of altered vesicle size and turnover remains unknown. But the functional consequence is measurable. Human iPSC-derived cardiomyocytes (hiPSC-CMs) pre-treated with wild-type EVs and then exposed to hydrogen peroxide (H2O2) showed only a 21% reduction in beat rate at 20 minutes. Cells incubated with Kcnma1⁻/⁻ EVs – or no EVs at all – showed an 82% reduction in beat rate. The miRNA mimics and inhibitors of miR-15a and miR-23a, reproduced the effect directionally, supporting the idea that differential miRNA packaging accounts for at least part of the functional differences. Moreover, EVs containing BKCa channels play an active role in protecting cardiac cells from oxidative stress.
Open Questions Regarding the Presence of BKCa in the EVs
One significant unknown is the orientation of BKCa in the EV membrane – specifically, whether the calcium-sensing domain faces the lumen or the exterior. This matters for understanding exactly when and how the channel activates during transit. Given that extracellular calcium concentrations are high (~1–2 mM), an externally-facing sensor would likely keep the channel constitutively active; a lumenally-facing sensor implies a different regulatory logic. The authors acknowledge this directly as a limitation.
The data are also derived from mouse-to-mouse cardiac injury experiments, so the leap to therapeutic application in humans requires substantially more work. What the study does establish – cleanly, with biophysical, proteomic, genetic, and functional evidence – is that EV membranes contain working ion channels, including BKCa, and that its presence shapes both the physical resilience of the vesicle and the biological potency of its cargo.

 (1097-1196) Antibody")
 Antibody")
