GLUT1, encoded by the SLC2A1 gene, is a membrane protein responsible for the facilitated diffusion of glucose across the plasma membrane. It belongs to the GLUT family of transporters and is widely expressed in many tissues, including erythrocytes, endothelial cells of the blood–brain barrier, and immune cells. GLUT1 ensures a basal supply of glucose required for cellular energy production and metabolic processes. Since glucose is a primary substrate for glycolysis and biosynthesis, GLUT1 is essential for maintaining cellular metabolism, especially in cells with high or rapidly changing energy demands.
GLUT1 is often described as a metabolic workhorse, but in adaptive immunity, it behaves more like a throttle. Two recent studies – one focused on B cells, the other on T cells – show that glucose influx through GLUT1 does not just sustain survival, but also quantitatively determines immune output. In one case, antibody quality and plasma cell (PC) generation depend on it, and in the other, effector cytokine production rises and falls with it.
Since GLUT1 activity depends on membrane localization, measuring its surface transporter levels is essential for interpreting metabolic capacity. This requires antibodies recognizing extracellular epitopes that allow flow cytometric detection without permeabilization. In both studies, surface GLUT1 expression was measured by flow cytometry – a method central to understanding the biology of GLUT1.
GLUT1 Controls Germinal Center (GC) Output in B Cells
In work from the University of Pennsylvania, Brookens and colleagues asked a direct question: once B cells have developed normally, do they still require sustained glucose influx through GLUT1 to mount an effective immune response (1)?
To answer this, they conditionally reduced the expression of murine Slc2a1, the gene encoding GLUT1, after establishing a normal pre-immune repertoire. The resulting phenotype was dramatic: early GC B cell numbers fell, PC output declined, and affinity maturation was impaired.
The mechanistic logic emerged from how glucose was being used. Reduced GLUT1 limited glucose entry into GC B cells. That, in turn, reduced flux into the pentose phosphate pathway (PPP). Redox buffering capacity dropped, and the anabolic support required for PC differentiation weakened. ATP production itself was not the dominant bottleneck. Instead, routing of glucose into the PPP and glycosylation pathways proved critical for sustaining antibody secretion and quality.
Importantly, GLUT1 expression was not inferred from transcriptional profiling of B cells. Instead, it was measured at the cell surface by flow cytometry using Alomone’s extracellular-directed anti-GLUT1 antibody conjugated to FITC (Anti-GLUT1 (extracellular)-FITC Antibody (#AGT-041-F)). That extracellular binding ability is important since transport capacity depends on membrane-localized transporter abundance, not total cellular protein levels. By quantifying surface GLUT1, the authors directly assessed the glucose import potential of GC B cells.
The research showed that in GC B cells, the glucose transport rate sets the ceiling for antibody quantity and quality. Surface GLUT1 expression in this case becomes a functional readout of metabolic competence.
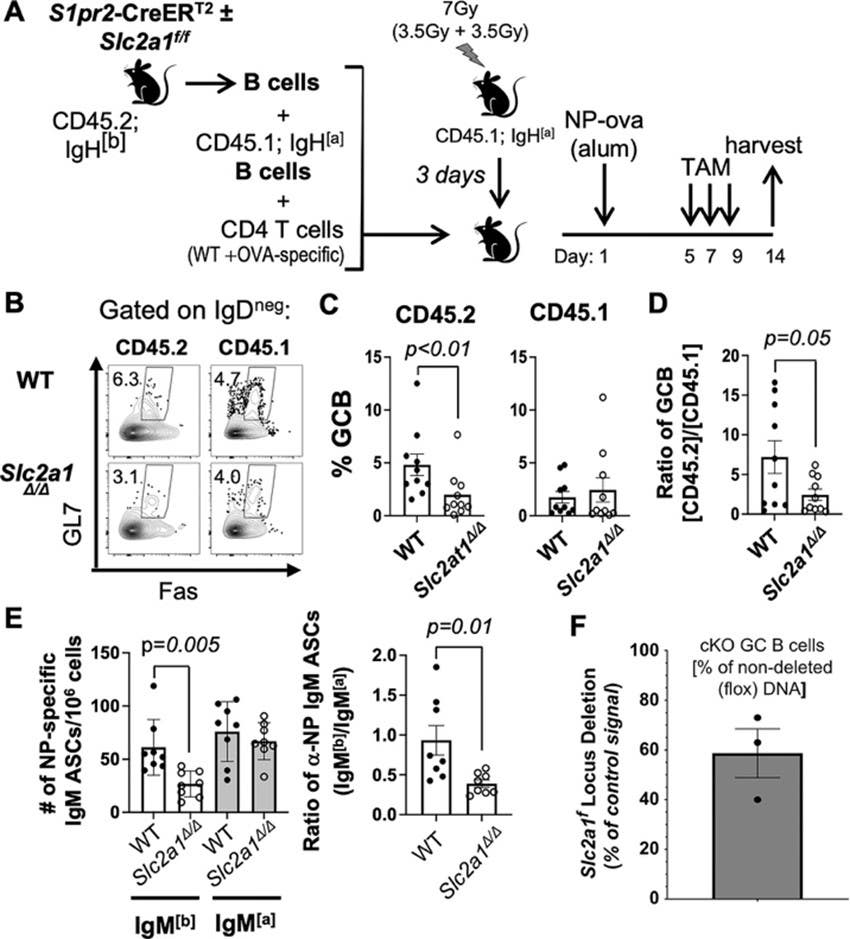
Figure 1. Glucose transporter 1 (GLUT1) expression within germinal center (GC) B cells regulates their prevalence and plasma cell (PC) generation. (A) Schematic model of adoptive transfer and immunization to test the effect of GC B cell-specific role of GLUT1. B cells from S1pr2-CreERT2 mice [wild-type (WT) – Slc2a1+/+ – or Slc2a1f/f] were mixed with CD45.1; IgH[a] B cells and CD4 T cells, then transferred into irradiated CD45.1; IgH[a] recipient mice followed by immunization with NP-ova. Starting 5 d thereafter, mice were treated with tamoxifen and harvested at day 14. (B-D) Flow plots (B), showed the quantified frequencies of Fas+ GL7+ cells in the B220+ IgDneg Dumpneg gate (Dump: CD11b, CD11c, F4/80, Gr1, 7-AAD) after gating to distinguish CD45.1 versus CD45.2 (C), and (D) the ratios of CD45.2 GCB / CD45.1 GC B cells are shown. (E) Numbers of all affinity NP-specific IgM[a] and IgM[b] ASCs that were derived from WT () and GCB-specific Slc2a1Δ/Δ (°) B cells (left), and the ratios of NP-specific IgM[b] ASC/ NP-specific IgM[a] ASC harvested from each recipient (right) (n = 8 WT S1pr2-CreERT2 B cell recipients and n = 8 mice that received GC-specific Slc2a1Δ/Δ B cells in three independent experiments). P values for the likelihood that the null hypothesis is correct were calculated by unpaired Student’s t-tests. (F) qPCR measurement of Slc2a1 floxed (flox) allele in freshly sorted CD19+ IgD− GL7+ CD38− GC B cells from tamoxifen-treated WT (S1pr2-CreERT2) and Glut1 cKO GCB (Slc2a1f/f; S1pr2-CreERT2) mice.
Image taken from Brokens et al. (2024) https://doi.org/10.4049/jimmunol.2200756.
GLUT1 Tunes Energetic Charge and IFN-γ Production in Effector T Cells
A complementary story unfolded in T cells from work by Chikina and colleagues at Institut Pasteur’s INSERM. They examined whether energetic charge – defined by the ATP:ADP ratio – varies across anatomical sites and time of day, and whether those differences influence effector function (2).
Their research showed that effector T cells displayed markedly higher ATP:ADP ratios in blood compared to lymph nodes. Energetic charge also peaked during the early rest phase in circadian cycles. At those moments of maximal energetic status, IFN-γ production was highest.
The mechanistic chain again points back to glucose transport. Effector T cells rely heavily on glycolysis, and when glucose availability rises, glycolytic flux increases, ATP:ADP ratios climb, and effector function intensifies. Naïve T cells, in contrast, did not exhibit the same spatial sensitivity, underscoring that differentiation state dictates the dependence on glucose transport.
Here too, GLUT1 surface expression was assessed by flow cytometry using anti-GLUT1 antibodies, including our Anti-GLUT1 (extracellular) Antibody (#AGT-041). The study reinforced a simple but often overlooked point: glucose uptake is limited by transporter abundance at the membrane. Measuring surface GLUT1 provides a mechanistic bridge between nutrient availability and functional output. The implication is that effector T cell function is dynamically tuned by nutrient access, with GLUT1 serving as the gatekeeper.
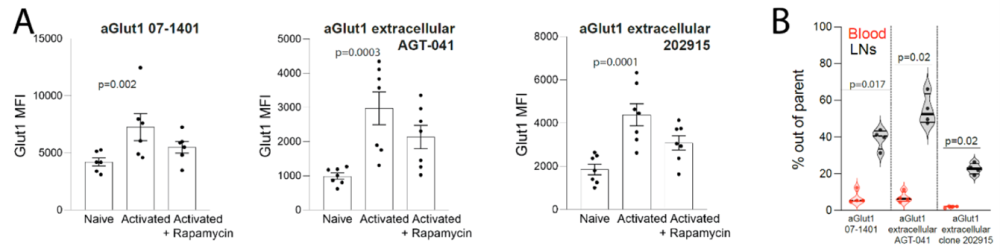
Figure 2. Detection of glucose transporter 1 (GLUT1)+ T cells assessed by three distinct anti-GLUT1 antibodies (Abs). A) OT-I T cells were activated with anti-CD3/CD28 Abs in the presence or absence of rapamycin or left unactivated. GLUT1 staining was performed using the three indicated anti-GLUT1 Abs, confirming activation-dependent GLUT1 expression. The data were compiled from 2 independent experiments with a total of 7 mice. B) The percentage of GLUT1+ cells among effector CD8+ T cells in the blood or lymph nodes measured with the three indicated anti-GLUT1 Abs. The data were compiled from 2 independent experiments with a total of 4 mice. Statistical analyses were performed using (A) Friedman and Dunn’s multiple-comparisons as well as (B) one-way ANOVA.
Image taken from Chikina et al. (2026). https://doi.org/10.1038/s41467-026-68559-1.
GLUT1: the Regulator
B cells and T cells use glucose differently, but both are constrained by transport. In GC B cells, GLUT1-dependent glucose influx supports anabolic routing through the PPP and glycosylation. This enables PC differentiation and high-quality antibody production. In effector T cells, GLUT1-associated glucose uptake sustains energetic charge and directly modulates cytokine output.
In both contexts, glucose transport is rate-limiting. It is not reliant on pathway selection or mitochondrial capacity, but the transport itself. This reframes GLUT1 from a metabolic marker to a quantitative regulator of adaptive immunity.
Why Surface Detection of GLUT1 Matters
GLUT1 trafficking is dynamic: activation signals alter its localization, nutrient conditions influence its stability, and total cellular GLUT1 does not necessarily reflect transport capacity, since uptake is constrained by membrane-localized transporters. This makes surface expression a very mechanistically meaningful readout.
For immunologists, antibodies directed against extracellular epitopes are indispensable. Surface-directed antibodies allow GLUT1 quantification without permeabilization, while preserving live-cell compatibility, and providing direct transport capacity in flow cytometry panels.
At Alomone Labs, we offer extracellular-directed anti-GLUT1 antibodies validated for flow cytometry. These antibodies are available in both unconjugated forms and forms directly conjugated to multiple fluorophores for multicolor panel integration. To our knowledge, we are uniquely positioned in combining extracellular targeting with several fluorophore-conjugated options for GLUT1.
In studies of immunometabolism, activation state, trafficking dynamics, or nutrient niche biology, surface GLUT1 measurement provides a mechanistically meaningful readout, since it links transporter abundance to immune output.


-FITC Antibody")
 Antibody")