
There’s a long-standing tension in enteric nervous system (ENS) biology between what genes say and what neurons do. Resolve that tension, and you often find something unexpected sitting in the middle. In a recent paper published in Purinergic Signalling, Hendler and colleagues (1) from Michigan State University and Sapienza University of Rome did exactly that – and what they found in the middle was a macrophage.
A Genomic-Functional Disconnect Worth Explaining
The enteric nervous system – the dense neural network embedded in the gut wall, sometimes called the ‘second brain’ – communicates extensively through purinergic signals. Adenosine diphosphate (ADP), generated by ectonucleotidase activity on released ATP, is a key mediator. In the myenteric plexus, enteric glia are highly responsive to ADP, mounting large calcium transients through P2Y1 receptors. Neurons, by contrast, seem comparatively indifferent to ADP despite single-cell sequencing data consistently showing that P2ry1 is actually more enriched in neurons than glia.
This kind of genomic-functional mismatch invites hypotheses. The one Hendler et al. pursued was elegant in its logic: P2Y12 receptors, which are Gi-coupled and thus inhibitory to calcium signaling, might be co-expressed in neurons – effectively silencing their P2Y1-mediated responses to ADP. The prediction followed naturally: block P2Y12, unmask the neurons.
The team combined live whole-mount immunohistochemistry with calcium imaging in transgenic mice expressing genetically encoded calcium reporters in neurons and glia. To map receptor expression with confidence, they used fluorescently conjugated antibodies targeting extracellular domains of both receptors – our Anti-P2Y1 Receptor (extracellular)-FITC Antibody (#APR-021-F) and Anti-P2Y12 Receptor (extracellular)-PE Antibody (#APR-020-PE) – applied to live tissue preparations of the mouse colon myenteric plexus. Live labelling with extracellular antibodies is useful here since it targets the functional pool of surface receptors rather than intracellular pools of uncertain relevance. That distinction, as it turned out, mattered enormously for the P2Y12 story.
The Surprise: P2Y12 Belongs to the Macrophages
P2Y1 labelling behaved as expected. Both enteric neurons and glia were positive, with expression in glial cell bodies and processes as well as neuronal somata – consistent with the genomic data. P2Y12, however, was a different story entirely.
Despite detectable P2ry12 transcript in published single-cell sequencing datasets for both neurons and glia, live immunolabelling found minimal P2Y12 protein in the myenteric plexus itself (Figure 1). Instead, robust P2Y12 expression appeared in the cells surrounding the ganglia – cells whose morphology and MHC-II co-expression identified them as muscularis macrophages.
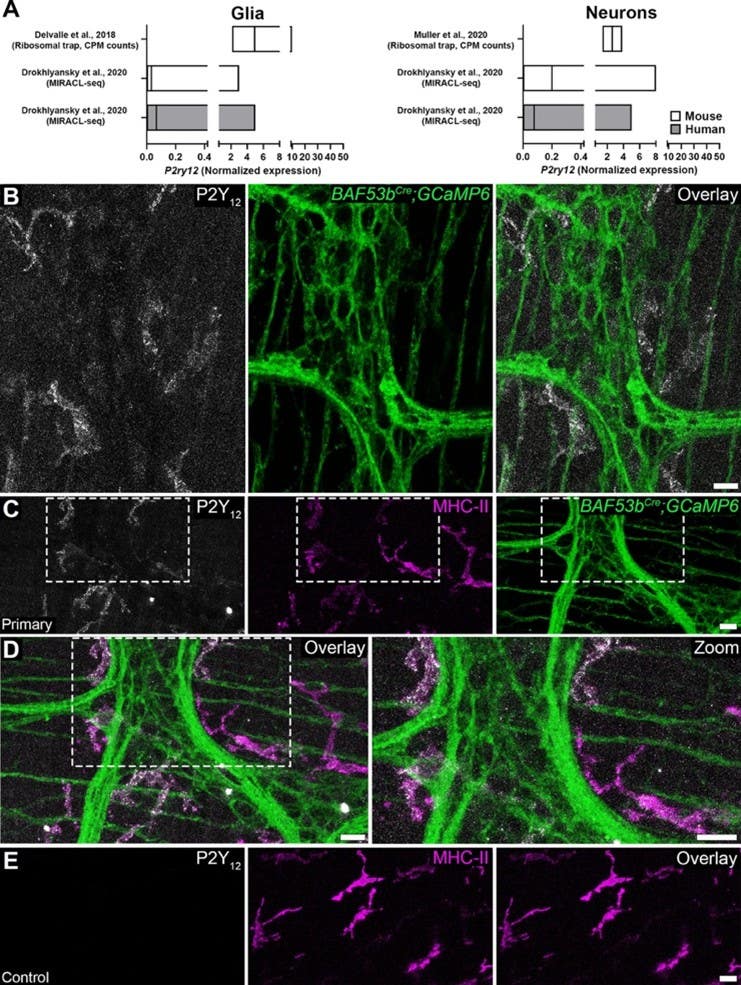
Figure 1. Cellular distribution of P2Y12 receptors in the colonic myenteric plexus. A P2ry12 cell-specific transcript expression in combined glial and neuronal subpopulations (left and right, respectively) from mouse and human colon prepared from published data sets. Graphs are prepared from the published datasets by [26, 35, 36] and data show normalized expression levels (mean ± range). B Representative confocal images showing P2Y12 immunoreactivity (grey) and GCaMP6f fluorescence (green) in neurons within the myenteric plexus of BAF53bCre; GCaMP6 mice. C-D Representative confocal images showing P2Y12 immunoreactivity (grey), major histocompatibility complex type II (MHC-II) immunoreactivity (magenta), and GCaMP6f fluorescence (green) in the myenteric plexus of BAF53bCre; GCaMP6. E Isotype control for P2Y12 antibody. Scale bars in B-E = 20 µm. n = 3 animals.
Figure from Hendler et al (2025). https://doi.org/10.1007/s11302-024-10060-9.
Further phenotyping revealed that the P2Y12-expressing macrophages were predominantly CD163⁺, a marker of tissue-resident, anti-inflammatory “M2-like” macrophages known to suppress enteric neurocircuit activity, in part by secreting bone morphogenetic protein 2 (BMP2). Of 137 cells quantified, 72.3% expressed P2Y12, with the majority co-expressing both CD163 and MHC-II. The receptor the team expected to find in neurons turned out to be the calling card of a resident immune cell with a known inhibitory role in ENS physiology.
Blocking P2Y12 Flips the Script on ENS Activity
To test the functional consequences, the team applied the selective P2Y12 antagonist PSB0739 to intact myenteric preparations and then rechallenged with ADP. The results were striking. Neuronal responses to bath-applied ADP increased in amplitude by roughly 33.5%, and total response area (AUC) by nearly 60%. Spontaneous neuronal activity doubled; spontaneous glial activity increased more than fourfold. Meanwhile, glial responses to ADP were substantially suppressed – nearly abolished in the bath application paradigm.
In short, blocking P2Y12 shifted ganglionic dynamics from glia-dominated, coordinated ADP responses to predominantly neuronal, stochastic firing. The organized, uniform glial response gave way to something considerably more chaotic.
What This Means for Gut Disease
The authors are careful about interpretation – and rightly so. The mechanism by which macrophage P2Y12 blockade suppresses glial activity is not yet clear, and exactly how macrophages translate P2Y12 signaling into inhibitory cues for neighboring neurons remains to be worked out. The source of constitutive ADP driving basal macrophage P2Y12 activity is an open question too, though a suggestive one: NTPDase2, which generates ADP from ATP, is almost exclusively expressed by enteric glia in the mouse colon. A signaling loop, perhaps.
But the broader implication is significant. As the authors put it: “Such mechanisms would be important to understand in more detail especially given that purines are major contributors to inflammatory signaling and changes in purinergic transmission between glia, neurons, and macrophages would have profound impacts on gut motor function.”
In conditions like inflammatory bowel disease or irritable bowel syndrome, shifts in macrophage phenotype are well documented. If CD163⁺ resident macrophages are the cells keeping ENS excitability in check through P2Y12 signaling, their loss or reprogramming during inflammation may be a direct contributor to the enteric hyperexcitability characteristic of these conditions. The fact that several widely used antiplatelet drugs – clopidogrel, prasugrel, ticagrelor – already target P2Y12 adds a layer of translational interest to these findings.
A New Aspect of Neuro-Immune Crosstalk
This study adds a functionally important piece to the growing picture of bidirectional communication between the immune system and the enteric nervous system. Muscularis macrophages have long been appreciated for their structural and trophic support of myenteric circuits; here they emerge as active purinergic regulators with the capacity to set the gain on neural excitability.
For ENS researchers, the lesson may be that understanding gut neural activity requires looking beyond the ganglion. The cells doing the most interesting signaling sometimes live just outside the frame.


-FITC Antibody")