
From sweet receptor cell turnover to insula-driven immune recall, new studies show the taste system is dynamically tuned by experience and internal state.
We tend to treat taste as a fixed input: sweet is sweet, bitter is bitter, and your tongue just reports what’s there. But that doesn’t hold up when you look closely. Taste operates far beyond simple flavor detection. It filters nutrient information, responds to context, and tracks prior exposure. Recent studies show that the taste system adjusts dynamically – across the tongue, in the supporting glial network, at nerve contact points, and deep into cortical circuits that link flavor to physiological state.
A high-sucrose diet shifts peripheral sweet responses. Local purinergic signaling between taste cells tunes sensitivity to sustained input. Gustatory nerves help maintain presynaptic structures needed for transmission. Moreover, in the brain, taste can retrieve immune memory through defined cortical loops. These findings reveal a sensory system that integrates diet, cellular signaling, synaptic maintenance, and learned associations.
In this latest instalment of our Senses series, we follow that progression – from altered taste receptor cell composition to circuit-level immune recall – each step showing how taste is continuously shaped by experience.
A High-Sucrose Diet Dulls Sweet Taste, but it’s Reversible
Short-term consumption of a high-sucrose diet selectively impairs peripheral sweet taste signaling in rats by dampening chorda tympani (CT) nerve responses and reducing sweet-responsive taste receptor cells. This occurs without affecting other taste modalities or structural features of the taste bud (1). Rats given a 30% sucrose solution for four weeks showed a dose-dependent reduction in CT responses to sucrose (0.15–2.0 M), while responses to NaCl, acids, umami, and thermal/tactile stimuli were unaffected.
Morphologically, sucrose-fed rats had no change in the number or size of fungiform taste buds on the front part of the tongue. In addition, no change in volume was observed when labeling P2X3⁺ (P2X3-positive) nerve fibers, which are sensory fibers that transmit taste signals; P2X3 is a purinergic receptor essential for ATP-mediated neurotransmission in the taste pathway. These P2X3+ nerve fibers were labeled with Alomone’s Anti-P2X3 Receptor Antibody (#APR-016). However, immunohistochemical staining revealed a significant drop in phospholipase C beta 2 (PLCβ2+) – a marker for type II taste cells (Figure 1), which are responsible for detecting sweet, umami, and bitter tastes. The effect was specific to the fungiform papillae and not observed for PKD2L1+ (type III marker) cells (that detect sour taste) or for general epithelial innervation.
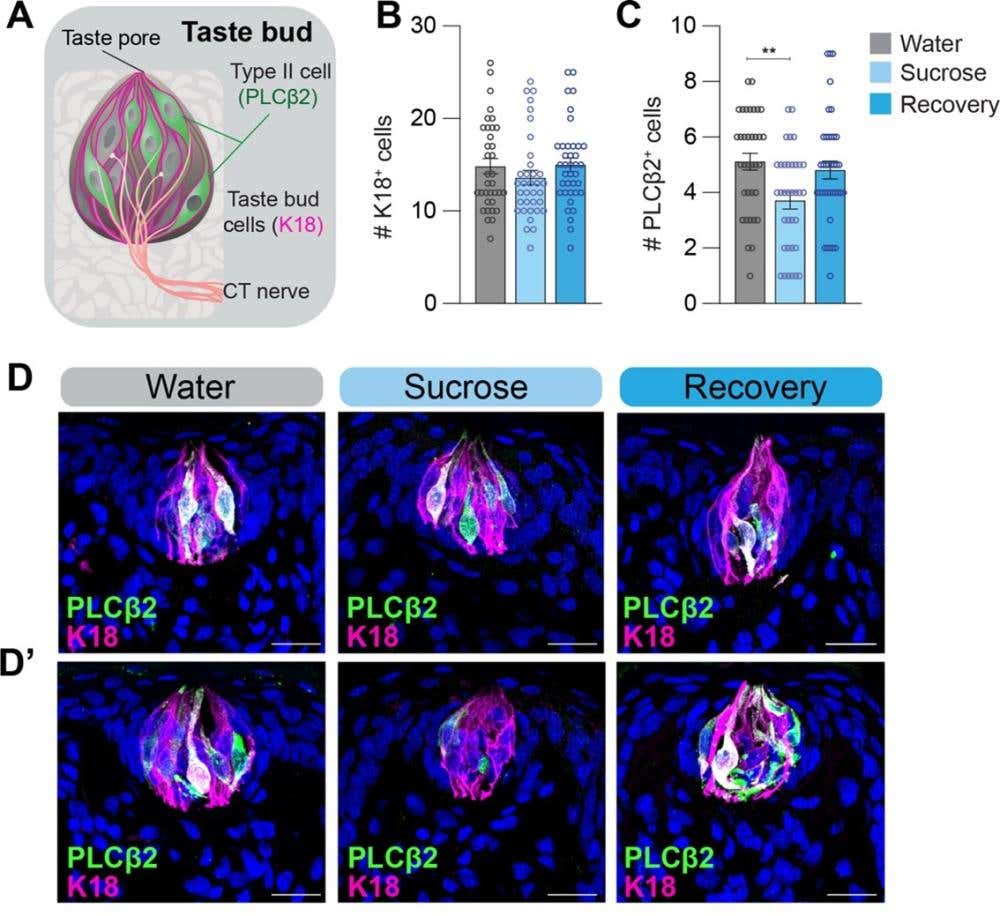
Figure 1. The number of phospholipase C beta 2 (PLCβ2)+ type II taste cells is lower in sucrose but not in recovery animals. A) Schematic of the taste bud (TB) with type II cells labeled with PLCβ2+ (green) and all taste cells labeled with keratin 18 (K18; magenta), which is a general marker for type I cells B-C) Quantification of the number of (B) K18⁺ cells and (C) type II PLCβ2+ cells in 10 μm tongue sections from three experimental groups: Water (control rats given only water), Sucrose (rats given a 30% sucrose solution for 4 weeks), and Recovery (rats previously exposed to sucrose, then returned to water for 4 weeks). n=6 tongues for each group; 4–8 TBs were analyzed from each tongue; each circle represents one taste bud. ** P ≤ 0.01. D-D’) Maximum intensity confocal projections of K18+ (magenta) and PLCβ2+ (green) cells from two representative rats per dietary condition (Water; Sucrose; Recovery). The scale bars represent 20 μm. Image adapted from Sung et al. (2022).
Switching animals back to water for four weeks restored both CT responses and PLCβ2+ cell counts, indicating that diet-induced taste changes are reversible and not due to permanent nerve loss or structural degradation.
Dietary sugar intake selectively and reversibly impairs peripheral sweet taste by altering receptor cell composition, not nerve input. These findings position peripheral taste plasticity as a dynamic, modality-specific response to dietary exposure – with implications for sensory regulation of food intake and metabolic health.
Glial Taste Cells Tune Sweetness Through P2RY2 Receptor Feedback
Changes in taste sensitivity are not limited to structural adaptation. Within taste buds, sweet responses are also dynamically tuned through intercellular signaling. Using intravital two-photon imaging, researchers showed that prolonged sweet stimulation activates type II taste cells, which release ATP that diffuses laterally to activate P2RY2 metabotropic receptors on adjacent type I (glia-like) cells (2). These type I cells, traditionally viewed as passive support, exhibited delayed but sustained calcium responses following sweet stimulation.
P2RY2 expression was localized to type I cells using our knockout-validated Anti-P2Y2 Receptor Antibody (#APR-010). P2RY2-deficient mice showed attenuated calcium responses in type I cells and delayed behavioral adaptation to sweet stimuli (longer latency to switch licking away from sweet solution during stimulus trains). Application of the P2RY2 agonist diquafosol enhanced adaptation, further confirming the role of this purinergic pathway in mediating taste adaptation dynamics.
Notably, the suppressive effect did not occur at the level of taste cell-neuron synapses. Instead, it occurred within the taste bud microcircuit, which suggested that glia-like cells act as a local inhibitory circuit that dampens prolonged sweet responses at the source.
Taste buds self-regulate sweet signaling through a glial feedback loop mediated by purinergic crosstalk. This mechanism provides fast, local control of stimulus persistence – reducing signal redundancy to the brain and allowing taste buds to maintain a dynamic range even during sustained stimulation.
Taste Receptor Synapses Depend on Nerve Signals
Structural and local plasticity both depend on intact innervation. In a mouse denervation model, researchers found that removing gustatory input from the CT caused a rapid and selective loss of presynaptic protein clusters in taste buds. This included the Bassoon scaffolding protein, and calcium homeostasis modulator 1 (CALHM1), the sweet, umami, and bitter channel, which mediates the neural transmission of these tastes (3). Over 90% of these presynaptic puncta were close to P2X2+ (P2X2-positive) nerve fibers under normal conditions [labeled using our knockout-validated Anti-P2X2 Receptor Antibody (#APR-003)], confirming tight spatial association with gustatory inputs (Figure 2).
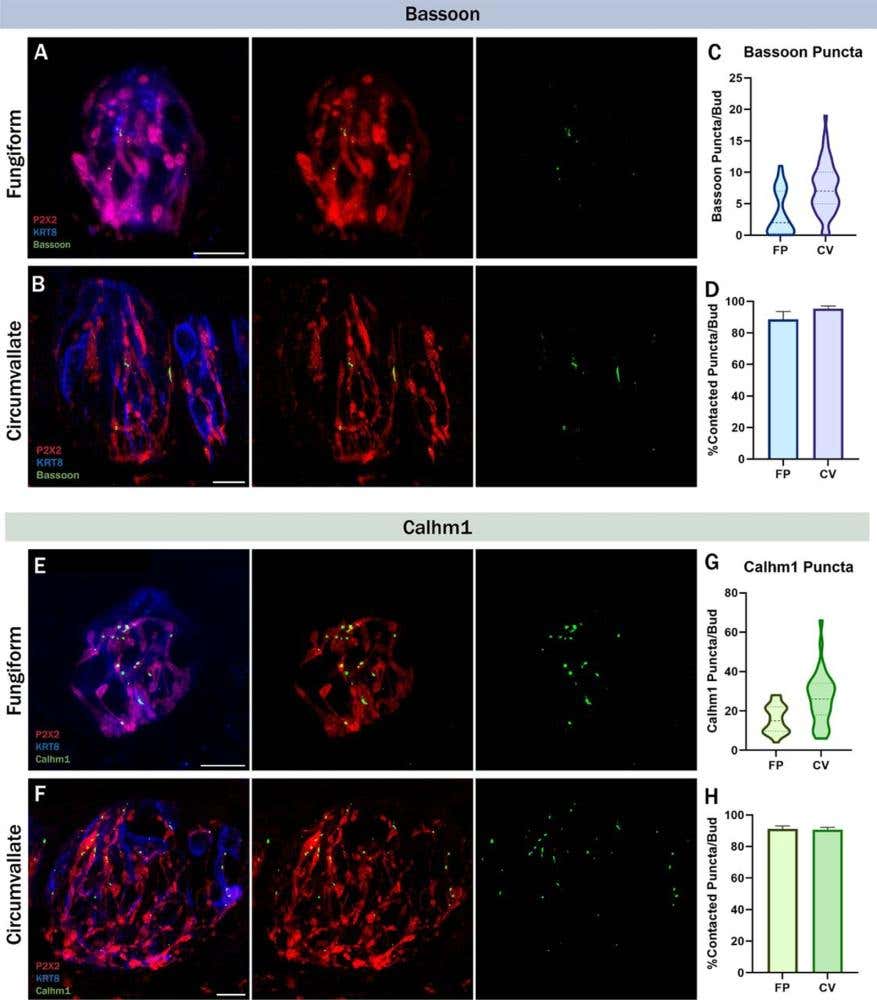
Figure 2. Bassoon and calcium homeostasis modulator 1 (CALHM1) are frequently contacted by gustatory neurons. Immunolabeling of P2X2 purinergic receptor (red), keratin 8 (KRT8; blue) a type II cell general marker, and Bassoon (green) in A) fungiform and B) circumvallate taste buds. C) Number of Bassoon puncta per taste bud in fungiform and circumvallate taste buds. D) Average percentage of Bassoon puncta that were contacted by P2X2-positive nerve fibers per taste bud. C,D) FP (Fungiform Papillae): n=36 buds/4 mice, CV (Circumvallate Papillae): n=57 buds/4 mice. Immunolabeling of P2X2 (red), KRT8 (blue), and CALHM1 (green) in E) fungiform and F) circumvallate taste buds. G) Number of CALHM1 puncta per taste bud in fungiform and circumvallate taste buds. H) Average percentage of CALHM1 puncta that were contacted by P2X2-positive nerve fibers per taste bud. G,H) FP: n=26 buds/4 mice, CV: n=39 buds/4 mice. The scale bar represents 10µm. Error bars: SEM. Image adapted from Landon et al. (2024).
Following complete nerve transection, Bassoon and CALHM1 expression dropped significantly within 3–5 days, even though the overall taste bud structure remained intact. The mRNA levels of both genes also declined, suggesting transcriptional downregulation in the absence of innervation. During subsequent reinnervation, presynaptic markers began to reappear – but only ~50% re-established contact with nerve fibers, and overall signal density remained below intact levels.
Presynaptic site maintenance in taste receptor cells requires trophic or contact-dependent cues from gustatory nerves. These results imply that gustatory neurons don’t just receive input from taste cells – they actively sustain their synaptic competence, possibly by delivering retrograde factors or stabilizing membrane specializations. Without that contact, taste buds lose their ability to signal, even if the cells themselves survive.
Taste Triggers Immune Recall via an Insula Brain Circuit
Peripheral modulation of taste shapes sensory encoding, but central processing determines what those signals mean. In male mice, the memory of a taste-immune pairing is stored in a bidirectional anterior-posterior insular cortex (aIC-pIC) circuit, enabling a learned immune response to be reactivated by taste alone (4). After pairing saccharin (a neutral taste) with IL-1β (an immunomodulatory stimulus), mice displayed both aversive behavior and increased splenic IFN-γ expression (a pro-inflammatory cytokine) in a delayed response. The same response occurred when re-exposed to saccharin alone, which indicated a conditioned immune response (CIR).
Circuit dissection showed that aIC-to-pIC projections were necessary for the behavioral (aversion) component, while bidirectional activity was required to retrieve the immune (IFN-γ) component. Chemogenetic inhibition of the aIC-pIC pathway blocked saccharin-induced aversion, while silencing both directions blocked the immune response entirely.
To confirm synaptic connectivity, the authors used Tetrodotoxin (#T-550) to block action potentials and CNQX (#C-140) to inhibit AMPA/kainate-mediated glutamatergic transmission, demonstrating that silencing both pathways completely abolished the immune response. These pharmacological tools helped confirm that aIC neurons directly excite pIC neurons through AMPA receptor-mediated fast glutamatergic transmission, and that this pathway is necessary for circuit activation during retrieval.
Taste cues can retrieve immune memory through a cortico-cortical loop that bridges sensory perception and immunological anticipation. This study shows that the brain can encode and later recall an immune state via sensory inputs – placing taste at the interface of experience-based regulation of immune readiness.
Taste is Dynamic – Shaped by Diet, Nerves, and Memory
Taste adapts to high dietary sugar by adjusting peripheral sensitivity. It suppresses signal overload locally through glial crosstalk. It depends on ongoing neural contact to maintain synaptic readiness. Furthermore, in the brain, it can recall past immune states through dedicated cortical circuits. Rather than passively reporting flavors, taste acts as a flexible gatekeeper, shaped by experience and tuned to physiological context.




