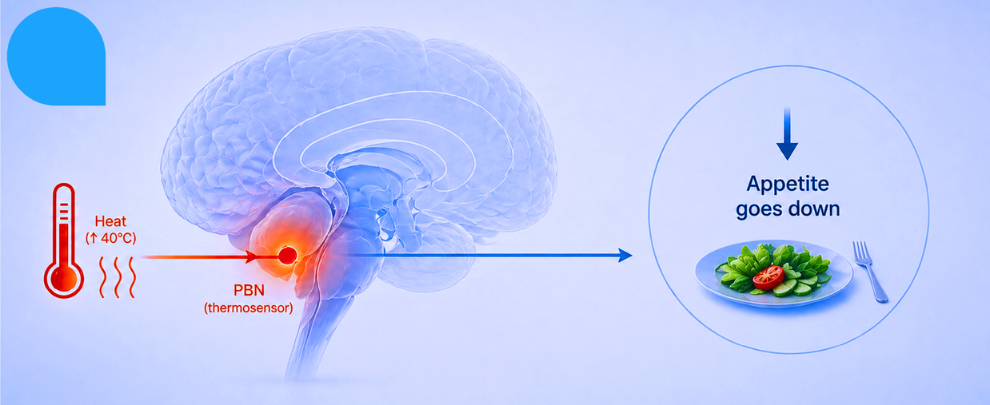
You probably already knew this – you’ve noticed it yourself: it’s 35°C outside, you’ve been on the bus for twenty minutes, you finally get home, and the last thing you want is a large bowl of pasta. A cold glass of water while reclining on the cool floor tiles, maybe. But pasta? No.
The observation that heat suppresses appetite is ancient. The mechanism behind it, though, has been sitting there largely unexplained – a phenomenological shrug filed under ‘seems obvious, probably metabolic.’ A new Nature paper from Benevento et al. decided to stop shrugging and actually map the circuit (1). What they found is architecturally elegant and involves a cell type that most people outside hypothalamic neuroscience have never heard of.
The short version is that heat activates neurons in the brainstem, those neurons connect to specialized glial cells in the hypothalamus, and those glial cells release a growth factor that silences hunger-promoting neurons, resulting in you eating less – for hours, or sometimes an entire day.
The Usual Suspects, and a Surprise
The parabrachial nucleus (PBN), tucked in the pons, is the brain’s primary thermosensor relay. It’s been well characterized; we know its glutamatergic neurons fire in response to heat, project to the hypothalamus, and perform various thermoregulatory functions. That part isn’t new. What’s new is what those projections are hitting.
Benevento et al. showed that glutamatergic PBN axons – VGLUT2+ ones, specifically – form synaptoid contacts directly onto α-tanycytes, a specialized ependymal cell type lining the wall of the third ventricle. Tanycytes have one soma touching the cerebrospinal fluid (CSF) and a long basal process extending deep into the hypothalamic parenchyma, terminating near blood vessels and neurons. They ferry signals in and out of the arcuate nucleus (ARC). And they are, depending on who you ask, the brain’s metabolic gatekeepers.
The authors used transsynaptic viral tracing (rAAV8-EF1a-mCherry-IRES-WGA-Cre injected into the lateral ventricle) to confirm that PBN neurons actually synaptically connect to tanycytes, not just pass nearby. They also used optogenetics [ChR2(H134R) expressed in VGLUT2+ PBN neurons, blue light pulses, and a patch-clamp on the tanycytes] and recorded excitatory postsynaptic currents (EPSCs) with amplitudes of around 7 pA and latencies of approximately 135 ms, with a failure rate of 77%. Modest, as direct synaptic inputs go. But functional.
More than 50% of the tanycytes received two or three VGLUT2+ terminals within 0.5 µm of their vimentin+ processes. α-tanycytes had a significantly higher density of inputs than β-tanycytes. And tanycytes expressed AMPA receptor subunits (confirmed using Alomone’s Guinea pig Anti-GluR1 (GluA1) (extracellular) Antibody (#AGC-004-GP) and Anti-GluR2 (GluA2) (extracellular) Antibody (#AGC-005)) with GluA2 preferentially expressed on α-tanycytes (Figure 1). Therefore, they have the molecular machinery to respond to glutamate. When action potentials are evoked in synaptically connected neurons, calcium transients are produced in tanycytes. If tanycyte AMPA receptors are blocked with NBQX, the transients disappear.
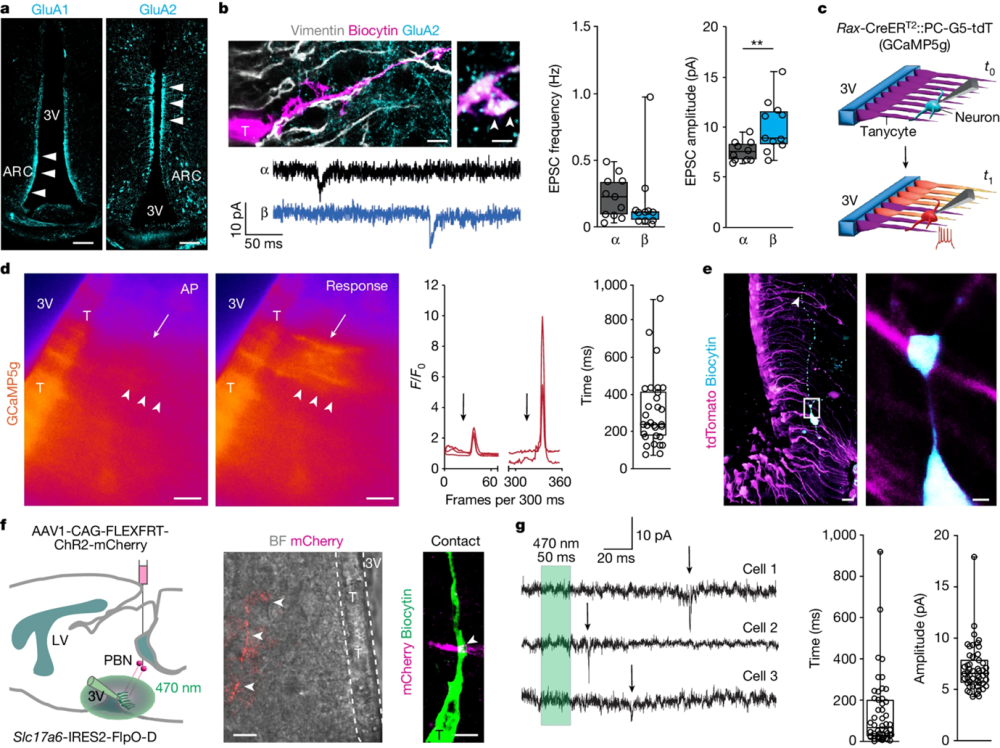
Figure 1. Tanycyte responses to synaptic afferent modulation. a, GluA1 and GluA2 in tanycytes. Scale bars, 100 µm. b, Top left, biocytin-filled, vimentin+ tanycyte (T) with GluA2 expression (main image; scale bar, 5 µm), with an enlarged view showing GluA2 in a vimentin+ basal filament (arrowheads; scale bar, 800 nm). Frequency (middle) and amplitude (right) of sEPSCs recorded in α- and β-tanycytes (both n = 11 cells). Detailed statistics are presented in the Methods. c, Schema of ex vivo experiments. Top, configuration at test (t0). Bottom, action potentials (APs) in neurons evoked at t1. d, Left, Ca2+ transients in tanycytes (arrowheads) in response to an evoked AP of 30 pA per 100 ms (arrow) (Supplementary Video 1). Middle, relative fluorescence intensity for GCaMP5g (F/F0) in tanycytes upon AP induction in neurons (arrows). Right, time lag of GCaMP5g relative fluorescence after the last AP (trains of 8 APs; 305.2 ± 35.25 ms, n = 30 tanycytes, n = 6 experiments). Scale bars, 20 µm. e, Left, a biocytin-filled neuron in the arcuate nucleus (ARC) of a Rax-CreERT2::PC-G5-tdT mouse. Right, intersection between a biocytin+ neuronal process (cyan) and a tanycyte (tdTomato+, magenta) in the outlined region in the left image. Scale bars: 20 µm (left); 2 µm (right). f, Left, cartoon showing tanycytes (green) tested for optogenetically induced EPSCs by stimulating pontine parabrachial nucleus (PBN) efferents (ChR2–mCherry, red) with 50-ms pulses of 470-nm light. Middle, bright-field (BF) view of tanycytes along the third ventricle overlaid on an mCherry+ afferent (arrowheads; scale bar, 10 µm). Right, a putative intersection between a tanycyte and afferent (scale bar, 2 µm). g, Left, optically induced EPSCs (arrows) in tanycytes. The time lag (middle; 256.5 ± 34.66 ms) and amplitude (right; 6.759 ± 0.48 pA; n = 29 EPSCs from n = 7 tanycytes, n = 4 independent experiments) are shown. b, d, and g, In box plots, the center line is the median, box edges delineate the top and bottom quartiles, whiskers extend to minimum and maximum values and circles depict individual data points.
Image taken from Beneveto et al. (2024). doi.org/10.1038/s41586-024-07232-3.
What Tanycytes Do with the Signal
This is where things get interesting. Exposure to 40°C for one hour induced significant cFOS expression in α-tanycytes in both male and female mice – 24 mice total across groups, with data replicated at two rostrocaudal positions in the ARC. Cold (4°C) did not produce this result, indicating that the tanycyte activation was heat-specific.
The authors interrogated single-cell RNA-sequencing data and landed on Vegfa as their candidate signaling molecule – tanycytes had it, and prior work had linked VEGFA to fasting-induced capillary remodeling in the ARC. Fluorescence in situ hybridization confirmed that 40°C exposure upregulated Vegfa mRNA in α-tanycytes. Moreover, chemogenetic activation of PBN glutamatergic neurons produced the same result without any temperature change at all. The PBN → tanycyte → VEGFA pathway is sufficient to trigger this, even at thermoneutrality.
A reasonable hypothesis at this point would be that VEGFA floods into the CSF for broad volumetric signaling. The authors tested this directly: they cannulated rats, exposed them to 40°C or 25°C for one hour, aspirated the CSF from the cerebellomedullary cistern, and ran an ELISA. They found no significant difference in CSF VEGFA. So that’s not it. Instead, VEGFA appears to be released locally along the basal processes of tanycytes, directly onto neurons in the ARC.
Silencing the Hungry Neurons
The ARC neurons in question are Flt1+ – FLT1 being the primary VEGFA receptor. Both AgRP+ and TH+ neurons (both orexigenic) expressed FLT1 in the ARC, and tanycyte processes were found in close apposition to their perikarya. POMC neurons – the “satiety neurons” – did not express FLT1 in this analysis, which provides the selectivity: VEGFA dampens the hunger-promoting side of the ARC, leaving the satiety-promoting side alone.
The electrophysiology added more data to top this. In ex vivo brain slices, increasing the superfusate temperature from 25°C to 38°C shifted the action potential threshold of ARC neurons from −33.66 ± 0.70 mV to −29.21 ± 1.98 mV (p < 0.001). That’s about 4.5 mV, but it’s enough to matter. Add axitinib, a VEGF receptor antagonist (40 µM), and the threshold shift disappears. The neurons don’t hypo-respond to heat; they respond to VEGFA released by tanycytes in response to heat. Remove the molecule, remove the effect.
The Circuit is Necessary and Sufficient
This is the part where the authors did the experiment you actually want them to do. They conducted two complementary manipulations:
First of all, they knocked down Vegfa in tanycytes using siRNA delivered to the third ventricle. The 40°C-induced reduction in food intake over 24 h was significantly attenuated. But the effect isn’t gone entirely – repeated ANOVA measurements showed a trend that falls short of significance in the VEGFA knockdown RNAi group – but it’s clearly blunted.
They blocked vesicular exocytosis from Rax+ tanycytes entirely using tetanus toxin light chain (TeLC) expressed in a Cre-dependent manner. Heat still activated PBN neurons normally – cFOS counts in PBN neurons were statistically identical between GFP controls and TeLC animals. But tanycytes go quiet (cFOS − throughout), and food intake is no longer suppressed by temperature switching (p < 0.01). Without tanycyte exocytosis, there’s no appetite suppression.
Secondly, they pursued the other direction by chemogenetically activating the glutamatergic PBN neurons at thermoneutrality and the animals ate significantly less. Next, they blocked tanycyte exocytosis first, then chemogenetically activated PBN again. The eating-time reduction disappeared. The PBN activation was necessary but not sufficient on its own – it required tanycytes to complete the circuit.
What This Actually Means
This paper described a discrete multi-node circuit in which an extrahypothalamic sensory relay – the PBN – links to a non-neuronal intermediary – α-tanycytes – to enforce a sustained metabolic response. The signal doesn’t stay in the CSF, but it also doesn’t diffuse broadly. Instead, it’s released focally along tanycyte basal processes onto specific ARC neurons, and it is selective for the orexigenic components of the circuit.
The practical implication for appetite biology is that there’s a glial node in the food intake circuit that can be modulated chemically and targeted by temperature – a finding with obvious relevance to thinking about metabolic disease, thermoregulation in aging populations, and why nobody ever wants a big lunch on a hot day.
The limitation to be aware of here is that this is a mouse model with controlled thermal manipulations. The 1°C-per-mouse variability in skin temperature and the predominantly male cohort in some experiments are real constraints. Whether this circuit operates identically in humans remains an open question. In fact, the human VEGFA-independent components of heat-induced anorexia remain unknown.
But in mice, the architecture is real, the mechanism is measured, and the cell nobody expected to be central to this story turns out to be doing quite a lot.

 (extracellular) Antibody")
 (extracellular) Antibody")