Loading...
Checkout
Login with a verification code
Enter your email address and we'll send you a verification code.
Code expires in
Checkout as a new customer
Creating an account has many benefits:
- See order and shipping status
- Track order history
- Check out faster
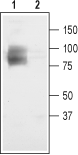
 Antibody")


 Antibody")
 Antibody")
