
Every sensory system begins with a conversion – light into voltage, vibration into current, chemicals into ion flux. But none is as intimate, or as universal, as touch.
The sense of touch starts with the simplest physical principle: force on a membrane. A hair bends, a cell stretches, a channel opens. That movement becomes charge, and charge becomes sensation. Yet simplicity hides depth. Touch isn’t one mechanism but rather it’s many overlapping layers – some mechanical, some chemical, some emotional. It persists through redundancy, adapts with age, and even links skin to reward.
This is the grand finale of our Senses series. After vision, hearing, taste, and smell, it’s time for the sense that never sleeps – a sense that can soothe or sting, warn or reward. Recent work has adjusted what we think about this sense, showing that touch is not a single pathway, but a system of channels and circuits that turn pressure into perception, and sometimes, into pleasure.
The Mole’s Star: Engineering for Contact
The star-nosed mole perceives the world almost entirely by touch. Each of its 22 nose rays is covered with Eimer’s organs – miniature mechanosensory domes packed with nerve endings. Instead of being defined by new genes, transcriptomic analysis of the mole’s trigeminal ganglia shows its neurons are instead a new balance of familiar ones. Mechanotransducers such as Piezo1, acid-sensing ion channel 3 (ASIC3), stomatin-like protein 3 (Stoml3), and the cyclic nucleotide–gated channel subunit A2 (Cnga2) were all enriched, while nociceptor markers declined (1). This shift turns a generalist sensory ganglion into a precision tactile interface.
Immunostaining using our anti-CNGA2 antibody (#APC-045) confirmed CNGA2 protein expression within the sensory fibers innervating Eimer’s organs and confirmed the transcriptome-based predictions of the role of CNGA2 in delicate touch (Figure 1). These findings reframed touch not as a unique innovation but as a reweighted use of conserved molecular parts. What differs between species – like us vs. the mole – is not the presence of touch genes, but how much real estate they occupy.
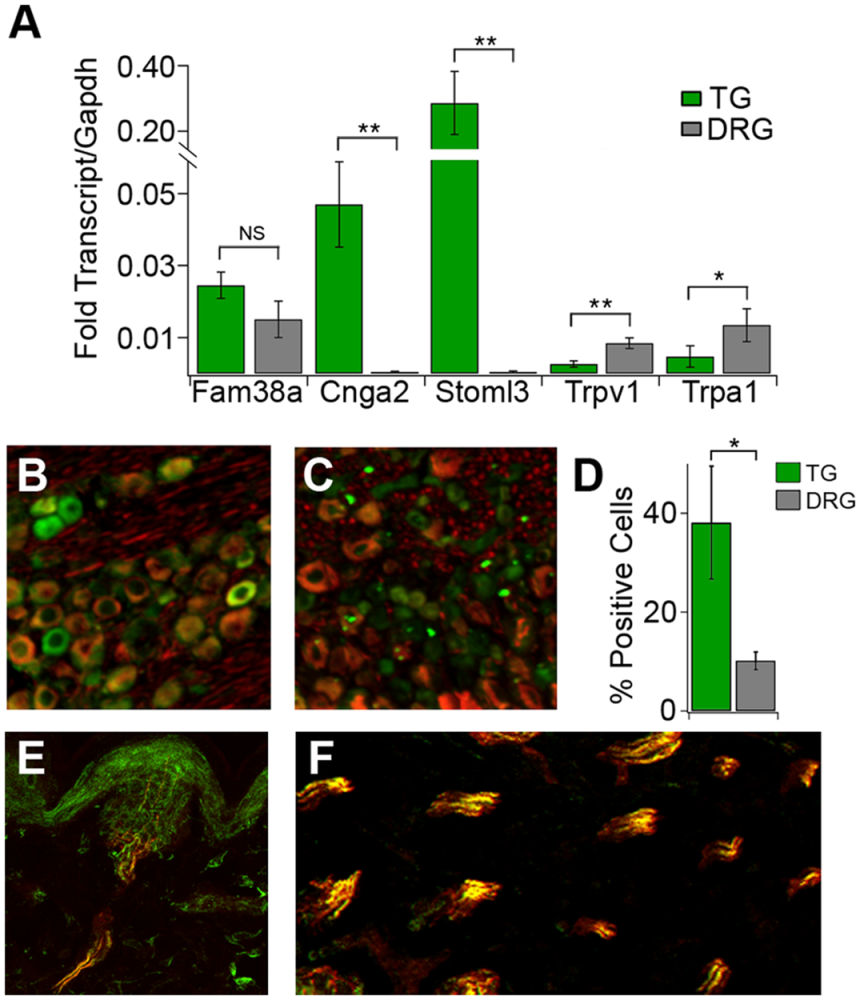
Figure 1. Genes linked to touch and pain show distinct expression patterns in the star-nosed mole. A) Gene expression analysis (qPCR) comparing the trigeminal ganglion (TG, green) and dorsal root ganglion (DRG, gray). The TG – which supplies the star organ – shows much higher levels of several touch-related genes, including cyclic nucleotide–gated channel subunit A2 (Cnga2) and stomatin-like protein 3 (Stoml3). Gene expression levels were normalized to Glyceraldehyde 3-Phosphate Dehydrogenase (Gapdh), a stable housekeeping gene commonly used as an internal reference (n = 3 samples per tissue). FAM38A= Family with Aligned Member 38A, influencing cell contractility and cytoskeletal organization. Trpv1= Transient Receptor Potential Vanilloid 1, plays a central role in pain sensation, inflammation, and thermosensation. Trpa1= Transient Receptor Potential Ankyrin 1, plays major roles in pain, itch and inflammation. B-C) Fluorescent staining for CNGA2 (green) together with NF200 (red), a marker for large touch fibers. Neurons in the TG show much stronger CNGA2 labeling (B) compared to those in the DRG (C). D) Quantification of CNGA2-positive cells confirms this difference: a much larger percentage of neurons express CNGA2 in the TG than in the DRG (n = 4 sections per tissue). E) A cross-section of a single Eimer’s organ showed strong CNGA2 staining (green) in the sensory nerve endings that penetrate this touch structure. F) Nerve bundles that innervate multiple Eimer’s organs also show robust CNGA2 expression [green, with substantial overlap with NF200-positive fibers (red)]. Asterisks mark statistical significance (**P < 0.01, *P < 0.05; NS = not significant, one-way ANOVA). Error bars represent S.E.M. Image taken from Gerhold et al. (2013), https://doi.org/10.1371/journal.pone.0055001. Licensed under Creative Commons Attribution 4.0 International (CC BY 4.0).
When Touch Becomes Pain
Mechanotransduction doesn’t only encode light contact; it can also encode discomfort. In a recent Fabry disease mouse model, loss of the enzyme α-galactosidase A (GLA) caused a buildup of glycosphingolipids (GS) that chronically sensitized the somatosensory system. Across the anterior cingulate cortex (ACC), spinal cord, and dorsal root ganglia, expression of the acid-sensing ion channel 1a (ASIC1a) climbed sharply with age – confirmed using our anti-ASIC1 antibody (#ASC-014) in a Western blot (Figure 2) (2).
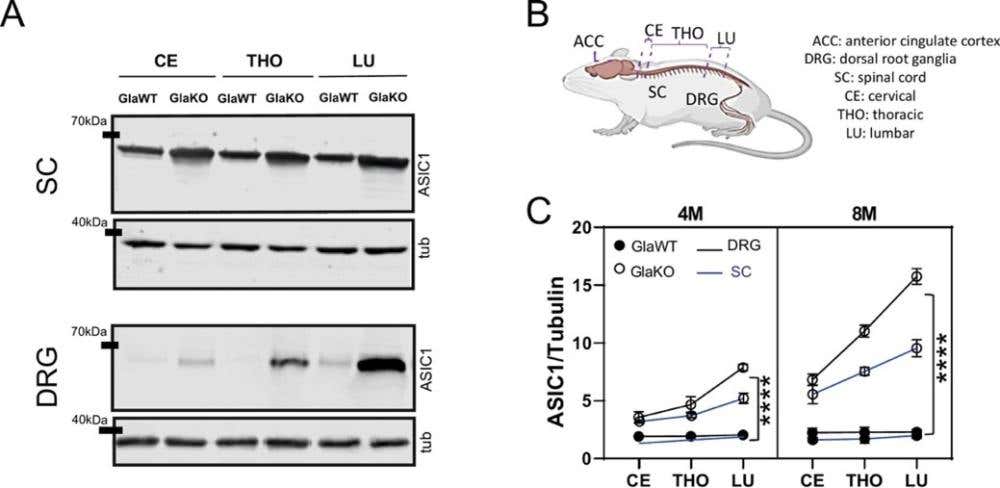
Figure 2. ASIC1 levels increase with age and throughout the pain pathway in α-galactosidase A knockout (GlaKO) male mice. A) Western blot images showing the amount of Acid-Sensing Ion Channel 1 (ASIC1) protein in different parts of the pain pathway. Samples were taken from the anterior cingulate cortex (ACC), the spinal cord (SC), and the dorsal root ganglia (DRG) in the cervical (CE), thoracic (THO), and lumbar (LU) regions of 4-month-old male mice. We compared wild-type mice (GlaWT) and mice lacking the Gla gene (GlaKO). Tubulin (tub) was used as a loading control. B) A simple diagram of the mouse nervous system highlighting the regions analyzed in the study. These include the ACC in the brain, the spinal cord, and the DRG at three levels—cervical, thoracic, and lumbar. C) Quantified levels of ASIC1 protein (normalized to tubulin) in 4-month-old (4M) and 8-month-old (8M) mice. In GlaKO mice, ASIC1 expression increases steadily from the cervical region down to the lumbar region in both the spinal cord (blue) and DRG (black). This upward trend becomes even more clear at 8 months of age. Each line connects measurements from the same tissue at different spinal levels to illustrate this gradient. The data are shown as mean ± S.E.M; ****P < 0.0001, n = 5 per group. Image taken from Montes et al. (2025), https://doi.org/10.3390/ph15121539. Licensed under Creative Commons Attribution 4.0 International (CC BY 4.0).
The overactive Acid-Sensing Ion Channel 1a (ASIC1a) drove extracellular signal-regulated kinase (ERK) phosphorylation and a striking mechanical hypersensitivity: animals recoiled to minor stimuli but barely responded to heat. When researchers blocked ASIC1a using our Psalmotoxin-1 (PcTx-1; #STP-200), mechanical thresholds normalized within minutes and phosphorylated ERK (pERK) levels fell back to baseline. The pain wasn’t an abstract outcome, but an electrical one, mediated by a defined ion pore.
This work linked metabolic disease to mechanosensory dysfunction and made a simple point: changing the local chemistry of the membrane – pH, lipid environment, ionic tone – can repurpose normal touch channels into chronic pain detectors.
What Happens When You Delete PIEZO2?
For a decade, mechanically activated ion channel 2 (PIEZO2) has been described as the master channel of touch and proprioception. Knockouts in mice lose fine tactile discrimination and position sense. But when researchers examined embryonic and early postnatal neurons lacking PIEZO2, something unexpected happened: mechanically activated currents remained (3).
Using an ultra-sensitive pillar deflection assay, the team measured nanometer-scale substrate motion in cultured dorsal root ganglion (DRG) neurons. In wild-type cells, about half responded with inward currents; in Piezo2-/- neurons, the proportion barely changed. Current amplitudes, thresholds, and adaptation profiles were almost identical, except for a moderate reduction in the fastest rapidly adapting events. The recordings included QX-314 (#Q-150) blocker to suppress voltage-gated sodium currents and isolate pure mechanically-gated responses. Even conditional knockouts tested a week after birth retained robust mechanosensitivity.
The result was a quiet revolution. PIEZO2 is crucial for behavior, but at the level of the cell, it’s not the sole gatekeeper of mechanical current. Other channels – possibly ELKIN1/TMEM87A, PIEZO1, or still-unknown families – may carry the load. Touch, it seems, is backed up.
When Touch Turns to Pleasure
Not all touch warns, guides, or informs – some rewards. A recent study mapped the skin-to-brain pathway that makes a caress feel good, tracing it to a rare class of sensory neurons known as mast cell-specific receptor for basic secretagogue 4 (Mrgprb4)-lineage fibers (4). The team used the AMPA receptor antagonist CNQX (#C-141) and the GABA(A) receptor blocker Bicuculline (#B-137) to characterize synaptic transmission between Mrgprb4-lineage afferents and spinal projection neurons.
When these neurons were optogenetically activated through the skin, female mice displayed a striking lordosis-like posture – a physical echo of sexual receptivity – and developed a strong preference for places where that stimulation occurred (Figure 3). Even in social isolation, activation of these neurons triggered dopamine release in the nucleus accumbens, the brain’s reward center.





-Bicuculline methochloride")