Gatekeepers of Tumor Metabolism and Therapeutic Targets
Cancer cells rewire metabolism by upregulating membrane transporters that enforce selective nutrient uptake. Not only do these transporters feed tumors, but they also impact redox state, signaling, and immune recognition. Vulnerabilities in these processes are now being targeted in clinical interventions.
To sustain their high metabolic demands due to unchecked growth and proliferation, cancer cells need a steady influx of nutrients, such as glucose, amino acids, fatty acids, and ions, often at levels that far exceed those required by normal cells. These demands are met by membrane transporters rather than passive diffusion, which act as selective gateways for metabolite entry and waste removal (1).
Among these, solute carrier (SLC) transporters play a central role. SLCs mediate the import of essential nutrients, support oncogenic signaling pathways, and allow tumors to adapt to hypoxia, acidosis, and nutrient scarcity. This dependency is not incidental. Many cancers upregulate specific transporters to meet the biosynthetic and energetic needs of their transformed state (2-4).
This altered transporter profile creates both a vulnerability and a biomarker. Overexpressed or dysregulated transporters offer potential points of therapeutic intervention. Some, like SLC1A5 or SLC7A5, are already being tested as drug targets or diagnostic markers. Others less characterized transporters , may hold value in specific tumor contexts, including breast cancer (3).
Here we will examine how transporters enable cancer metabolism and discuss which transporter families are crucial for this process. Finally, we describe how researchers are starting to exploit this biology for therapeutic gain.
The Basics: What Are Transporters?
Transporters are membrane-bound proteins that control the movement of molecules into and out of cells. They regulate access to nutrients, maintain ion gradients, remove waste, and coordinate signaling across cellular compartments.
When it comes to nutrient uptake and drug resistance, two transporter families are particularly important: the ATP-binding cassette (ABC) transporters and the SLC transporters (5). ABC transporters use ATP hydrolysis to export drugs, lipids, and xenobiotics; they are best known for their role in multidrug resistance. In contrast, SLC transporters mostly rely on ion gradients and facilitated diffusion to import a wide range of metabolites – including glucose, amino acids, fatty acids, vitamins, and metal ions – making them crucial for cancer metabolism (6).
There are over 400 human SLC genes grouped into more than 65 families. Many SLCs are highly tissue-specific, regulated by nutrient status or signaling pathways, and dysregulated in disease. In cancer, tumor cells frequently hijack or overexpress key SLCs to meet altered metabolic demands. For example, SLC2A1 [a.k.a. glucose transporter 1 (GLUT1)] imports glucose to fuel glycolysis, while SLC1A5 [a.k.a. alanine-serine-cysteine transporter 2 (ASCT2)] and SLC7A5 [a.k.a. L-type amino acid transporter (LAT1)] mediate amino acid uptak. This uptake is essential for activating mTOR – a central regulator of cell growth, metabolism, proliferation, survival, and redox balance (3, 4).
As well as being nutrient gateways, transporters also affect signaling cascades, drug resistance, and immune visibility. Their surface location and substrate specificity make them attractive candidates for targeted therapies and companion diagnostics.
Metabolic Requirements of Cancer – Why Transporters Matter
Tumors rewire cellular metabolism to support rapid growth, survival under stress, and immune evasion. This reprogramming includes a shift toward aerobic glycolysis, known as the Warburg effect, in which cancer cells preferentially convert glucose to lactate even in the presence of oxygen (7). But the Warburg effect is only one part of the story. Tumors also increase the uptake of amino acids, lipids, and micronutrients to meet biosynthetic demand.
These inputs are not passively acquired. Transporters are essential for supplying the metabolites that fuel oncogenic programs. Overexpression of SLC transporters enables tumors to import glutamine, leucine, serine, as well as other substrates needed for nucleotide synthesis, redox control, and epigenetic modification (8).
For example, SLC1A5 (ASCT2) supports glutamine uptake in many tumor types (9). Once inside the cell, glutamine serves the following functions: (1) it acts as a carbon and nitrogen source for the tricarboxylic acid (TCA) cycle, (2) it supports NADPH production for redox homeostasis, and (3) it regulates mTORC1 (an mTOR complex) activation via intracellular exchange with essential amino acids. SLC7A5 (LAT1) performs a related function by importing branched-chain amino acids such as leucine, which also promote mTOR signaling and protein synthesis (10).
Increased transporter expression is often associated with therapy resistance. Some tumors upregulate glucose transporters following chemotherapy, while others shift reliance to alternate amino acid carriers when targeted with inhibitors. This metabolic flexibility makes transporters not only drivers of malignancy but also exploitable vulnerabilities that can be targeted with drugs or combination therapies.
Spotlight: Transporters and Breast Cancer
Breast cancer subtypes differ widely in metabolic phenotype, with aggressive forms like triple-negative breast cancer (TNBC) showing increased reliance on nutrient uptake pathways (11). Several SLC transporters are overexpressed in breast tumors, where they support proliferation, survival, and therapy resistance (Table 1). Their expression patterns often correlate with hormone receptor status, proliferation markers, and treatment outcomes (12).
SLC7A5 (LAT1)
SLC7A5 is upregulated across multiple cancer subtypes including luminal, human epidermal growth factor receptor 2 positive (HER2+), and TNBC. SLC7A5 imports essential amino acids such as leucine in exchange for glutamine. In addition, SLC7A5 expression promotes mTOR activation, cell cycle progression, and resistance to endocrine therapy (13).
SLC6A14
SLC6A14 is a sodium- and chloride-dependent transporter with broad substrate specificity. It is highly expressed in estrogen receptor–positive (ER⁺) and TNBC tumors. SLC6A14 supports amino acid influx for biosynthesis of proteins and energy production. SLC6A14 has also been linked to tamoxifen resistance (14).
SLC38A1 (SNAT1)
SLC38A1 is elevated in high-proliferative tumors, particularly those with activation of the PI3K (phosphoinositide 3-kinase)–Akt (Protein Kinase B) signaling pathway, a key driver of cell growth, survival, and metabolism. SLC38A1supports glutamine uptake and contributes to redox balance and growth signaling (15).
SLC1A5 (ASCT2)
SLC1A5 is overexpressed in HER2+ and TNBC tumors. SLC1A5 facilitates glutamine import, fuels mitochondrial metabolism, and supports mTORC1 activation (9).
These transporters represent both biomarkers and potential therapeutic entry points for subtype-specific intervention.
Table 1. Major SLC transporters implicated in breast cancer.
| Transporter | Subtype/Context | Mechanism/Role in Cancer |
| SLC7A5 | Most subtypes | mTOR activation, amino acid exchange, growth |
| SLC6A14 | ER⁺ and TNBC | Amino acid influx, resistance to tamoxifen |
| SLC38A1 | High-proliferative tumors | Glutamine uptake, Akt pathway activation |
| SLC1A5 | HER2+, TNBC | Glutamine import, mTORC1 signaling |
Therapeutic Targeting: Current Approaches and Clinical Implications
The overexpression and functional requirement of transporters in cancer make them attractive drug targets. Unlike intracellular enzymes, many transporters reside on the plasma membrane and have defined substrate pockets – offering both accessibility and selectivity.
Small-Molecule Inhibitors
Several small-molecule inhibitors targeting SLC transporters have entered preclinical and early clinical testing:
- SLC7A5 (LAT1): JPH203, a selective LAT1 inhibitor, has shown antitumor activity in multiple solid tumors, including breast and thyroid cancer. It reduces mTORC1 signaling and amino acid influx and is currently in phase I/II trials (16).
- SLC2A1 (GLUT1): Inhibitors such as BAY-876 and WZB117 impair glycolysis and tumor growth in preclinical models. Selectivity remains a challenge due to GLUT1 expression in normal tissues (17).
- SLC16A1 (MCT1): While not discussed in earlier sections, MCTs are essential for lactate export in glycolytic tumors. AZD3965 (MCT1 inhibitor) has entered clinical trials for lymphoma and solid tumors with high lactate output (18, 19).
Antibodies
Antibody-based inhibition of transporters is less common but offers advantages in selectivity and prolonged half-life. Candidates targeting extracellular regions of ASCT2 or CD98 (a.k.a. SLC3A2, the chaperone for LAT1/2/xCT) are under development, with preclinical data showing reduced nutrient uptake and decreased tumor cell viability (20).
RNA-Based Approaches
siRNA and antisense oligonucleotides targeting transporter mRNA (e.g., SLC6A14 or SLC1A5) have shown efficacy in reducing tumor growth in animal models (21, 22). These strategies remain limited by delivery challenges but are gaining traction in combination approaches.
Combination Therapies
Monotherapies targeting transporters often have limited efficacy due to redundant pathways. As a result, most therapeutic strategies now focus on combination therapy:
- Co-targeting GLUT1 and ASCT2 to block both carbon and nitrogen sources (23).
- Pairing LAT1 inhibitors with mTOR inhibitors to block downstream signaling (24).
- Combining transporter blockade with immune checkpoint inhibitors, aiming to sensitize metabolically rigid tumors to immune attack (25).
Transporters also influence drug uptake and resistance. For example, ABC transporters export chemotherapy agents and can be inhibited to restore drug sensitivity. Meanwhile, some SLCs affect drug pharmacokinetics by altering cellular uptake of nucleoside analogs or tyrosine kinase inhibitors.
Studying Transporters: Antibodies and Research Reagents
Targeting transporters in cancer requires therapeutic candidates alongside validated reagents to study their expression, localization, and function. Antibodies against transporter proteins are essential for immunohistochemistry (IHC), flow cytometry, western blotting, and live-cell assays – especially when distinguishing intracellular vs. extracellular epitopes or for comparison across species (Table 2).
Binding Orientation Matters
Antibodies targeting extracellular domains are valuable for live-cell labeling, flow cytometry, and development of potential therapeutics. Intracellular domain antibodies, on the other hand, are preferred for fixed-cell analysis, functional evaluation, and mechanistic studies. For example:
- SLC2A1 (GLUT1) antibodies are available in both intra- and extracellular-binding forms, useful for IHC or in vivo blocking studies.
- SLC7A5 (LAT1) and SLC7A11 (xCT) antibodies are available for both intracellular tracking and surface detection depending on the research requirement.
Species Reactivity
Antibody reactivity with human, mouse, or rabbit proteins is critical for translational research. Mouse reactive antibodies enable tumor studies in xenograft models, while human-specific reagents are essential for validating clinical relevance.
Table 2. Selected antibodies to target transporters in cancer. Extracellular/cell surface-binding antibodies are labelled as “extracellular.” Conjugation refers to the following type of fluorophore the antibody is conjugated to: fluorescein isothiocyanate (FITC), R-phycoerythrin (PE), allophycocyanin (APC), or mFluor Violet™ 450. For help building a panel, you can use our Panel Builder – simply choose your targets, pick your antibodies, and it maps out the best possible fluorophore combinations for your setup. Reactivity refers to mouse (M), rabbit (R), or human (H).
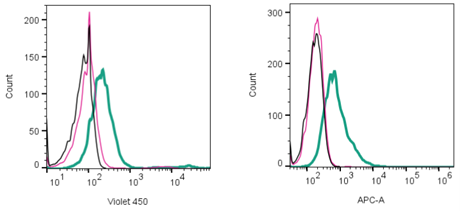
Figure 1. Cell surface detection of proteins by direct flow cytometry. Left: xCT/SLC7A11 detection in live intact human U-87 MG glioblastoma cells. Black) Cells. Red) Cells + Rabbit IgG isotype control-mFluor™ Violet 450 (#RIC-001-V). Green) Cells + Anti-xCT/SLC7A11 (extracellular)- mFluor™ Violet 450 Antibody (#ANT-111-V), (2.5µg). Right: APC (Allophycocyanin fluorophore) detection in live intact human U-87 MG glioblastoma cells. Black) Cells. Red) Rabbit IgG Isotype control-APC (#RIC-001-APC). Green) Cells + Anti-MCT1 (SLC16A1) (extracellular)-APC Antibody (#AMT-011-APC), (2.5μg).
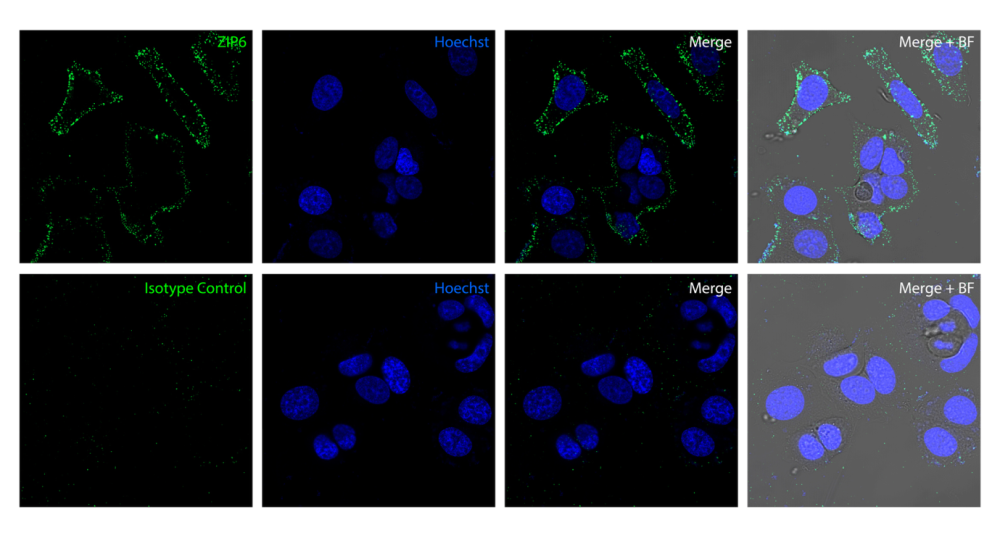
Figure 2. Detection of ZIP6 in human MCF7 cells. Top: ZIP6 was detected in live human breast adenocarcinoma MCF7 cells by confocal microscopy using Anti-ZIP6/SLC39A6 (extracellular) Antibody (#AZT-006, 1:25), followed by goat anti-rabbit secondary antibody conjugated to Alexa Fluor® 488 (green). The nuclei were stained with Hoechst 33342 (blue). Bottom: Isotype control staining was performed using Rabbit IgG Isotype Control (#RIC-001, 1:25), followed by goat anti-rabbit secondary antibody conjugated to Alexa Fluor® 488 (green). BF = Bright-field microscopy.
Validated antibodies with known epitope locations and defined cross-reactivity are essential for experimental reproducibility. You can find the epitope sequence listed on our data sheets when looking for your antibody. Many of these reagents are already in use for tumor panel screening, transporter knockout validation, and drug uptake studies.
Challenges and Future Directions
Despite growing interest in transporter biology, several barriers limit their clinical translation. Chief among these is redundancy. Many nutrients can be imported by multiple SLC family members, often with overlapping substrate specificity. For example, when ASCT2 (SLC1A5) is inhibited, some tumors upregulate LAT1 (SLC7A5) or SNAT2 (SLC38A2) to maintain glutamine flux (4). This redundancy blunts the efficacy of single-agent therapies and demands combination strategies or upstream interventions that affect transporter expression.
Another challenge is tumor heterogeneity. Transporter expression varies not only between cancer types, but also between patients, within tumors, and over time. Some transporters, like GLUT1 or LAT1, show broad overexpression. Others, like SLC6A14 or xCT, are enriched in specific subtypes or stress conditions. Without molecular stratification, clinical trials targeting transporters risk failure due to patient mismatch.
Compensatory metabolic rewiring is another limitation. Tumors can switch fuel sources – substituting glucose for acetate, or glutamine for serine – depending on transporter availability, oncogenic signaling, and microenvironmental pressures (26, 27). This plasticity undermines the long-term success of transporter-targeted monotherapies.
Future success will depend on a few key directions:
- Biomarker-driven patient selection: identifying tumors with high transporter dependence – through transcriptomics, imaging [e.g., fluorodeoxyglucose positron emission tomography (FDG-PET)], or immunoassays – will be essential for stratifying trials.
- Combination approaches: co-targeting multiple transporters, or combining transporter inhibition with mTOR inhibitors, immunotherapy, or chemotherapy, may overcome metabolic escape routes.
- Transporter–immune interactions: emerging evidence suggests that nutrient transporters shape the tumor microenvironment and influence T cell function, particularly through amino acid competition and redox regulation. These links offer opportunities to modulate immune visibility and resistance indirectly (3).
Finally, improved in vivo models and antibody-based approaches will help clarify transporter roles across tissues, disease stages, and therapeutic contexts.
Conclusion
Transporters are not passive players in cancer biology. They are dynamic gatekeepers of nutrient flow, modulators of intracellular signaling, and contributors to therapy resistance. SLC transporters, in particular, support the metabolic reprogramming that distinguishes tumors from normal tissues.
Data from multiple studies confirm that cancer cells upregulate specific SLCs – such as GLUT1, ASCT2, LAT1, and xCT – to meet the demands of uncontrolled proliferation and stress survival. This altered transporter expression creates measurable vulnerabilities. Several inhibitors are now in preclinical and clinical evaluation, while validated antibody tools allow for spatial and functional mapping across tumor types.
In breast cancer, transporter profiles vary by subtype, with distinct dependencies in HER2+, TNBC, and hormone receptor–positive tumors. These differences underscore the need for stratified therapeutic approaches and functional diagnostics.
As research advances, transporter targeting is likely to intersect with immunotherapy, metabolic imaging, and resistance management. Future success will rely on identifying where transporter dependence is an essential part of biology and matching the right tools to the right context.


-mFluor™ Violet 450 Antibody")

 (extracellular)-APC Antibody")
 Antibody")
