Control Products

BDNF Blocking Peptide (#BLP-NT010)
40 µg
$297.00
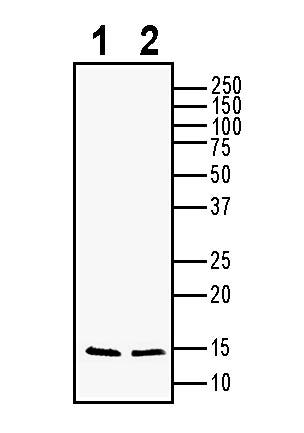
BDNF Positive Control for Western Blot (#PCP-B250)
1 unit
$245.00
Related Products
Mouse Anti-Rat p75 NGF Receptor (extracellular) Antibody
 Antibody")
Mouse Anti-Rat p75 NGF Receptor (extracellular) Antibody
View Product


Anti-p75 NGF Receptor (extracellular) Antibody
 Antibody")
Anti-p75 NGF Receptor (extracellular) Antibody
View Product
Anti-p75 NGF Receptor (extracellular)-ATTO Fluor-550 Antibody
-ATTO Fluor-550 Antibody")
Anti-p75 NGF Receptor (extracellular)-ATTO Fluor-550 Antibody
View Product

 Antibody")
Anti-TrkB (extracellular)-ATTO Fluor-488 Antibody
-ATTO Fluor-488 Antibody")
Anti-TrkB (extracellular)-ATTO Fluor-488 Antibody
View Product
-FITC Antibody")




Recombinant human BDNF proDomain (Val66Met) protein
 protein")
Recombinant human BDNF proDomain (Val66Met) protein
View Product
