
Deleting neurons and treating pain like a molecular scalpel – resiniferatoxin (RTX) is something you should definitely be looking at.
RTX, which is derived from the cactus-like plant, Euphorbia resinifera, is powerful stuff. It’s so powerful in fact that rather than just being mildly annoying like its 1,000-fold less potent analog, capsaicin, RTX holds vanilloid receptor subtype 1 (TRPV1) ion channels wide open to allow sodium and calcium to flood in. Leave it long enough and the intracellular calcium load becomes so catastrophic that it wipes out the nerve ending entirely.
Take a quick look at our RTX data in cells. You can see just how well RTX induces a calcium influx via TRPV1 activation at concentrations as low as the nanomolar range:
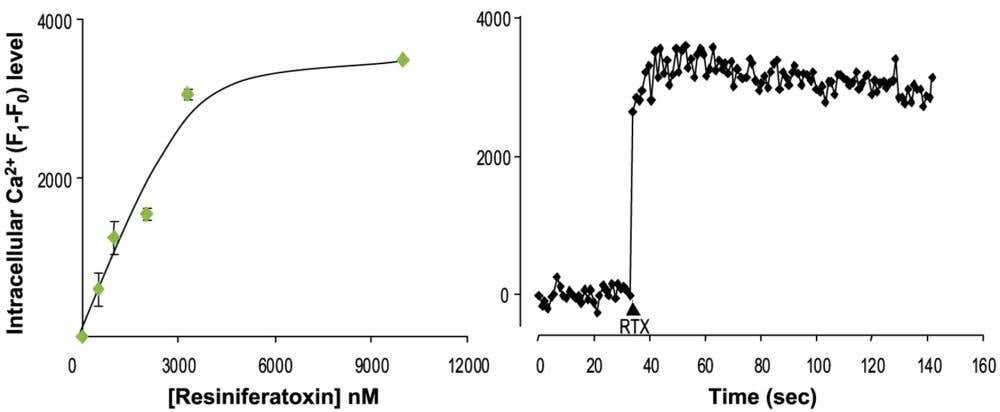
Figure 1. Resiniferatoxin (RTX) triggers a rapid, dose-dependent Ca²⁺ influx in TRPV1-expressing HEK293 cells. These graphs show how HEK293 cells engineered to express the TRPV1 channel respond to resiniferatoxin. Using the calcium-sensitive dye fluo-3 AM, intracellular Ca²⁺ changes are presented as ΔF (F₁–F₀), where F₀ is the baseline fluorescence before stimulation and F₁ is the fluorescence after stimulation, essentially reflecting the increase in calcium levels over baseline levels. In the left panel, escalating concentrations of Alomone’s Resiniferatoxin (#R-400) produce a clear, dose-dependent rise in ΔF (ED₅₀ ≈ 2 µM). In the right panel, the time-course plot shows that adding 10 µM of resiniferatoxin leads to a rapid and sustained increase in intracellular Ca²⁺, highlighting robust TRPV1 activation. The data were generated by the Alomone Labs R&D team.
But unlike general neurolytic agents, RTX is choosy. It goes after TRPV1-expressing nociceptors specifically – the Aδ and C-fiber population that actually carries pain. This is why people started calling RTX “molecular neurosurgery” and why the phrase “molecular scalpel” (1) is only slightly hyperbolic.
A Molecular Scalpel
Suddenly, you have a very potent chemical with a high affinity for afferent sensory pain neurons that leaves other sensory neurons unaffected. That should pique your interest as RTX has real potential in the realms of relieving pain in a specific area.
By injecting RTX at a discrete location, it physically deletes TRPV1-expressing Aδ and C-fiber cells in the dorsal root ganglia (DRG) (2). When you eliminate the sensory neurons, you effectively stop pain transmission along the spinal cord and up to the brain before it begins – all the while leaving touch, pressure, and proprioception untouched.
Fixing Knee Pain and More
The pain-relieving benefit of RTX has already been explored in canine research where the pain relief lasted for months to even years (3) – much more than the expected 2–3 weeks. The authors hypothesize that in addition to shutting off nociceptive neural activity, RTX may have disease-modifying actions. How? By affecting inflammation states. Since neurogenic inflammation at least partly maintains chronic inflammation levels, RTX may lessen this chronic inflammation, with subsequent reductions in plasma extravasation and edema.
Interestingly, while we’re talking about inflammation, a team from Germany used Alomone’s Resiniferatoxin (#R-400) and Anti-TRPV1 (VR1) Antibody (#ACC-030) to show that intrathecal RTX blunts TNF-induced pain behavior in rats (4). As illustrated in Figure 2, silencing of TRPV1-positive DRG neurons shifts the inflammatory tone: less TNF signaling, less edema, less neurogenic flare. Fewer reasons for the joint to complain. The far-reaching effects of RTX continue to be impressive – not to mention raising lots of new questions to pursue.
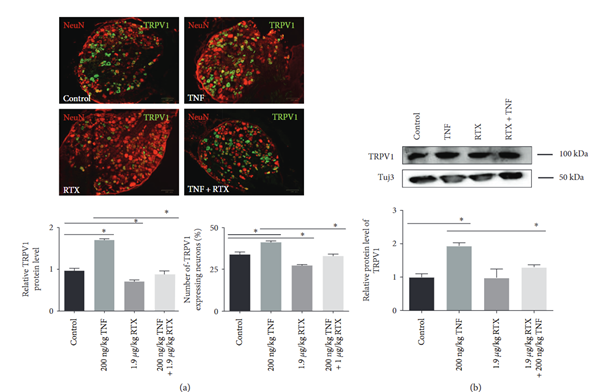
Figure 2. TNF factor increases TRPV1 receptor expression in sensory neurons, while resiniferatoxin (RTX) reverses this effect. A) Representative images showing dorsal root ganglion (DRG) neurons from adult rats stained for the NeuN neuronal marker (red) and TRPV1 (green) 24 hours after different treatments. TNF strongly increases TRPV1 signal intensity and the number of TRPV1-positive neurons. RTX treatment alone decreases TRPV1 expression, and when TNF and RTX are given together, RTX prevents the TNF-induced rise in TRPV1 levels. The bar graphs show the quantification of these effects, which revealed changes in TRPV1 protein levels and in the proportion of TRPV1-expressing neurons under each condition. B) Western blot analysis confirms these findings at the protein level: TNF elevates TRPV1 expression, whereas coadministration of RTX suppresses this increase. Image was taken from Leo et al. 2017.
More recently, we’re already seen preliminary trials of RTX treatment in humans to determine its efficacy in ameliorating localized pain in diseases such as knee osteoarthritis (5). A 2025 clinical review goes further and pulls the whole story together across canine and human osteoarthritis (6). The picture is pretty consistent: inject RTX into an osteoarthritic knee, defunctionalize the TRPV1 nociceptor terminals at the joint, and you achieve robust, long-lasting analgesia from a single injection. The effects often last up to six months and in some cases close to a year, while sparing motor function and proprioception. Since the treatment deletes the entire pain-sensing TRPV1 nerve ending, and does not just block a single receptor, the authors suggest that RTX also inhibits neurogenic inflammation at its source and may create a more favorable environment for tissue remodeling rather than just dulling symptoms.
RTX in Human Trials: Turning Off Pain at the Source
Treating knees may seem quite specialized, so what about broader pain issues? Patients with bone metastases or advanced abdominal tumors have widespread pain and often end up drowning in opioids. RTX offers a different route: ablate the nociceptors driving that agony.
The NIH team led by John Heiss delivered intrathecal RTX directly into the cerebrospinal fluid. The first cohort in their early phase I study was administered 3–26 µg under deep sedation, because yes, RTX itself is spectacularly painful on the way in (7).
The latest update: 19 patients, single intrathecal dose, followed for six months (8). Safety was the primary outcome, and in an end-stage oncology population you expect plenty of serious adverse events. Even so, RTX still managed to cut “worst pain” scores by ~38% and reduce opioid use by 57% at day 15. A few patients lost heat sensitivity in the relevant dermatomes, which is exactly what you’d expect if RTX did its job.
It’s not gentle. But for people with intractable end-of-life pain, RTX looks like one of the few tools that can actually break the circuit.
RTX Rewires Pain Pathways
Genomic analyses comparing RTX and capsaicin showed that while RTX hits harder, it also produces a cleaner transcriptional footprint. It downregulates several pain-associated genes (Camk2a, Scn11a, Adcy5, Cacna2d1, Ntrk1, Runx1, and Scn10a) more effectively than capsaicin (9).
It also suppresses “undesired” pain-related genes such as Kcnk2, Kcnj5, Gal, Nt5e, Gfra2, and Ptgdr – a broader shutdown of nociceptive machinery. That level of transcriptional pruning probably contributes to its long-lasting analgesia. It also reminds us that, yes, there are bound to be side effects we still haven’t mapped. So, while RTX is undoubtedly more potent than capsaicin (with a far more complex chemical structure, as you can see in Figure 3), there remains the potential for RTX to have unknown side effects.
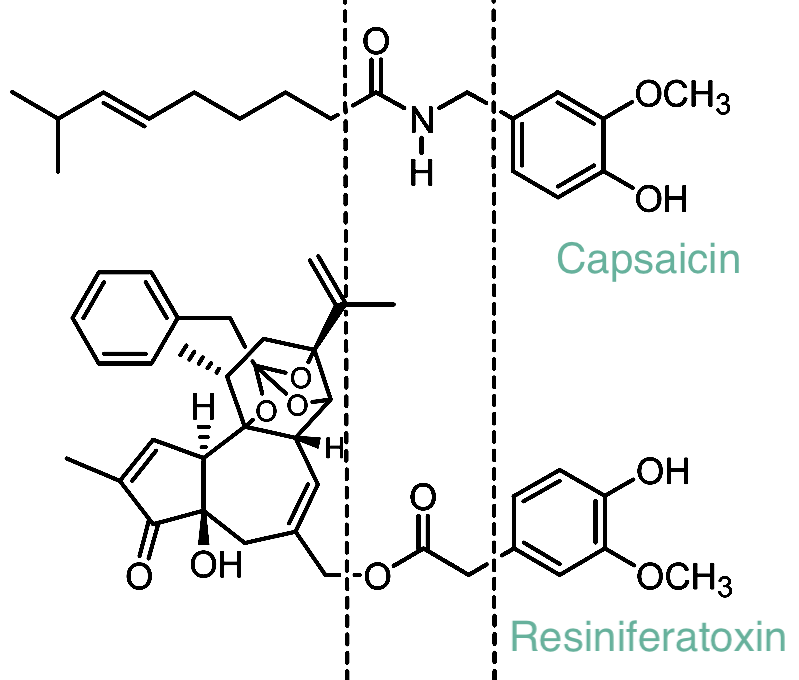
Figure 3. Structural comparison of capsaicin and resiniferatoxin. This illustration highlights the shared vanilloid “head group” found in both capsaicin and resiniferatoxin (in the middle). The head group allows for the activation of the TRPV1 receptor. While capsaicin has a relatively simple linear structure, resiniferatoxin contains an expanded, highly rigid diterpene core. This added structural complexity makes resiniferatoxin a far more potent TRPV1 agonist, producing dramatically stronger and longer-lasting activation compared to capsaicin.
RTX in Mechanistic Ion Channel Work
RTX is roughly 1,000-fold more potent than capsaicin. And unlike capsaicin, which causes transient excitation, RTX tends to leapfrog straight to desensitization and terminal deletion (10). Which is why a lot of labs pair the two: use capsaicin as a fast, noisy hammer, and RTX as the long-term switch you flip when you really want to shut a pathway down.
But RTX isn’t just for killing neurons – it’s also a favorite tool in ion channel biophysics, especially in groups exploring TRPV1 gating. Two recent papers show just how cleanly RTX stabilizes the open state (11, 12):
- RTX binding contributes ~6.8–7.4 kcal/mol of free energy toward TRPV1 activation.
- Each subunit contributes roughly equally, which fits the classic allosteric Monod-Wyman-Changeux (MWC) model surprisingly well.
- Using Alomone’s Resiniferatoxin (#R-400) and 6’-Iodoresiniferatoxin (#I-805), researchers built concatemeric TRPV1 channels with defined numbers of functional binding sites. This let them measure how channels behave with 0–4 RTX molecules bound – something that’s essentially impossible with normal ligand assays.
The outcome? TRPV1 behaves very close to an ideal allosteric protein, with only marginal deviations depending on which subunits carry the ligand. Those deviations are tiny – around 0.3–0.4 kcal/mol – and much smaller than the overall activation energy contributed by RTX binding. In other words: RTX is a superb probe for dissecting TRPV1 gating mechanics.
And since RTX binds so tightly, our RTX shows up constantly in TRPV1 electrophysiology papers. When someone needs a ligand that won’t fall off the channel mid-recording, RTX is the natural choice.
Blocking TRPV1 Instead of Blowing it Open
RTX has a sibling worth mentioning: iodo-resiniferatoxin (I-RTX), an iodinated form of RTX. It uses the same scaffolding but behaves differently. Instead of opening TRPV1, it blocks it – forcefully (13):
- I-RTX binds TRPV1 with low-nanomolar affinity
- It induces no TRPV1 current on its own, even at 10 µM
- It completely blocks capsaicin-evoked currents with an IC₅₀ = 3.9 nM
- In mice, it prevents capsaicin-evoked pain at doses as low as 16 ng (intrathecal)
It’s at least 40-fold more potent than capsazepine, the old-school antagonist. We supply both RTX and I-RTX [available as either 5’-Iodoresiniferatoxin (#I-800) or 6’-Iodoresiniferatoxin (#I-805)], so you can push TRPV1 in either direction – open it until the neuron dies, or block it until it goes quiet.
Beyond its headline trick of shutting down TRPV1 without triggering the fiery mess that capsaicin gives you, I-RTX has a few other perks worth knowing about. It’s a genuinely clean antagonist – not a partial agonist in disguise – so you don’t get weird baseline currents or flare responses. Since it binds hard and fast, it doubles as a high-affinity probe for mapping TRPV1 density in tissues, and unlike capsazepine it barely touches other TRP channels. It blocks heat activation as well as ligand activation, it keeps capsaicin desensitization artifacts from ruining your data, and it behaves impressively predictably in vivo. If you want TRPV1 to become inactivated without detonating the neuron, I-RTX is the molecule.
The Takeaway
RTX is one of those molecules that looks absurd on paper and somehow goes even further in the lab. It kills the right neurons, rewires pain pathways, modulates inflammation, and behaves beautifully in mechanistic ion channel studies. And depending on how you deliver it, you get transient agonism, long-term silencing, or full-on molecular ablation.
If you’re working anywhere near nociception, inflammation, TRP channels, or sensory neuron biology, you should at least have RTX, I-RTX, or a TRPV1 antibody in your freezer. We make all of these in-house, and yes, they’re as potent as the papers make them sound. If your project needs something even more specific, we can also create custom-made variants tailored to your research –so you get exactly the tool you need to ask the right biological questions.




 Antibody")

