
In 2025, the cell surface became the place where decisions were made.
Across oncology, metabolism, and neuroscience, many of the most interesting papers of 2025, all published in first-tier journals, had the same quiet protagonist: the cell surface. Receptors, ion channels, and cytoskeletal nanostructures did more than just relay information – they decided which cells invade, which starve, which are expelled, and which survive.
Come along for a guided tour through the work published in 2025, where we look at how cells sense their world and turn those signals into fate, decisions, and behavior.
When Receptors Decide Cell Fate
Melanoma researchers targeted a part of the neurotrophin receptor typically ignored: the transmembrane domain (TMD). Screening >8,000 small molecules against the p75NTR TMD identified compounds that physically wedged into this helix. Receptor-specific engagement was validated using an Anti-p75 NGF Receptor (extracellular) Antibody (#ANT-007) and medicinal chemistry refinement produced the compound Np75-4A22. This molecule bound directly to the p75NTR TMD, blocked NGF-induced recruitment of fascin, and prevented filopodia formation in human melanoma cells at sub-micromolar concentrations (1).
Disrupting this single membrane segment had organism-level consequences. Orally delivered Np75-4A22 showed high bioavailability, low toxicity, and significantly reduced melanoma lung invasion in mice. This study established that receptor TMDs can be used as actionable drug targets in metastatic disease.
In the lung, large conductance Ca2+– and voltage-activated K⁺ (BK) channels functioned as anti-inflammatory brakes. In lipopolysaccharide (LPS)-induced pneumonia, pharmacological BK activation with two unrelated agonists, NS-1619 (#N-105) and NS19504 (#N-235) (2) reduced neutrophil infiltration, CCL-2 chemokine levels, reactive oxygen species, and H₂O₂, while increasing superoxide dismutase and catalase. Therefore, these agonists reduced lung inflammation and oxidative stress, while enhancing antioxidant defenses. These effects were reproduced in primary human alveolar epithelial cells and reversed by the BK blocker Paxilline (#P-450), confirming channel specificity. Together, agonist-antagonist pairing positioned BK channels as druggable suppressors of inflammatory lung injury.
In the heart, capillary pericytes emerged as pressure-sensitive amplifiers of ischemic damage. In cultured cardiac pericytes, mechanical stress increased cytosolic Ca2+ through Transient Receptor Potential Melastatin 4 (TRPM4) and TRPM7 channels, an effect abolished by channel inhibitors (3). Channel expression and localization were confirmed using our Anti-TRPM4 Antibody (#ACC-044) and Anti-TRPM7 Antibody (#ACC-047) (Figure 1). In vivo, TRPM4 inhibition reduced the infarct size by ~3.5-fold in a rodent acute myocardial infarction (AMI) model, linking pericyte mechanotransduction directly to ischemic outcome.
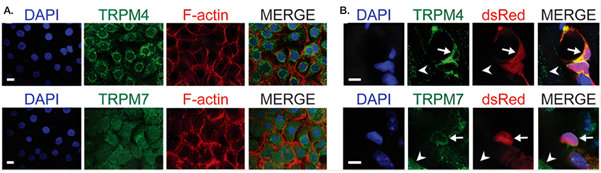
Figure 1. TRPM4 and TRPM7 are expressed in primary cardiac pericytes. Representative fluorescence images showing the expression of the ion channels TRPM4 and TRPM7 in cardiac pericytes. A) In cultured primary cardiac pericytes, cell nuclei were labeled with DAPI (blue), the actin cytoskeleton with F-actin (red), and TRPM4 (upper panel) or TRPM7 (lower panel) with green fluorescence. The merged images illustrate cellular localization: TRPM4 is enriched in perinuclear structures, whereas TRPM7 displayed a more diffuse, membrane-associated distribution. B) In heart tissue sections from transgenic mice expressing DsRed (red) specifically in pericytes, TRPM4 (upper panel) and TRPM7 (lower panel) signals co-localized with DsRed-positive cells in the coronary vasculature (arrows), consistent with pericyte identity. Occasional staining in non-vascular cells was also observed (arrowheads). The scale bars represent 20 µm (cultured cells) and 5 µm (tissue sections).
Image adapted from Methner et al. (2025), https://doi.org/10.14814/phy2.70396. Licensed under Creative Commons Attribution 4.0 International (CC BY 4.0).
At epithelial surfaces, high cell density alone acted as a selective pressure, favoring cells that could tolerate crowded conditions. The density activated epithelial sodium channel (ENaC), depolarized cells through Na⁺ influx. Cells with sufficient ATP repolarized via the Na⁺/K⁺-ATPase, while ATP-poor cells remained depolarized (4). Persistent depolarization activated the voltage-gated potassium channels (KV) 1.1 and KV1.2, as well as the volume-regulated anion channel LRRC8A/SWELL1, driving K⁺, Cl⁻, and water efflux. Using Alomone’s Anti-KV1.1 (KCNA1) (extracellular) Antibody (#APC-161), Anti-KV1.2 (KCNA2) Antibody (#APC-010), and Anti-LRRC8A (extracellular) Antibody (#AAC-001) to map this extrusion, researchers showed that this ionic cascade induced cell shrinkage and triggered live-cell extrusion. Blocking any component disrupted this extrusion, identifying an energy-sensitive membrane module that selectively removes metabolically weak cells.
Across melanoma, lung endothelium, cardiac pericytes, and epithelia, remarkably small differences in membrane-mediated control of ions, voltage, and mechanical force tipped cells past points of no return, irreversibly determining their fate.
Where Force Meets Function
Osmosensory neurons resolved subtle volume changes using nanoscale membrane architecture. In these cells a truncated form of the transient receptor potential vanilloid subtype 1 (ΔN-TRPV1) ion channel was activated by physical forces inside the cell. These channels are located in small gaps in the actin cytoskeleton, where mechanical forces can easily reach and activate them (5). During hypertonic stress, these sites exhibited amplified membrane displacement. Proximity ligation assays using Anti-TRPV1 (VR1) Antibody (#ACC-030) and β-tubulin antibody showed close association of ΔN-TRPV1 with cytoskeletal anchoring points that experienced exaggerated mechanical strain. Osmosensitivity therefore depended not only on channel properties, but on cytoskeletal geometry.
A parallel principle appeared in the auditory brainstem. At the endbulb of Held synapse, monoaminergic varicosities containing norepinephrine (NE) and serotonin (5-HT) were anatomically positioned near glutamatergic terminals (6). Immunostaining relied on multiple 5-HT receptor antibodies, including those in the Serotonin Receptor Antibody Explorer Kit (#AK-585). Functionally, NE – but not 5-HT – increased the frequency of miniature excitatory postsynaptic currents (mEPSCs), without affecting evoked transmission or neuronal excitability. mEPSCs are tiny, spontaneous synaptic currents, providing a readout of background synaptic activity at excitatory synapses. Here, neuromodulation adjusted synaptic noise rather than signal fidelity.
When Peripheral Sensors Decide What We Eat, Taste, and Seek
Membrane sensors also shaped behavior. After catabolic stress, mice avoided protein rather than seeking it, even when total caloric intake dropped (7). This aversion was traced to glutamine, lysine, and threonine metabolism in the gut, which generated ammonia sensed by Transient Receptor Potential Ankyrin 1 (TRPA1) on enterochromaffin cells. TRPA1 activation triggered serotonin release and brainstem signaling that suppressed protein intake. TRPA1 expression was mapped using Anti-TRPA1 (extracellular) Antibody (#ACC-037). Genetic or pharmacological disruption of TRPA1 or serotonergic signaling abolished protein avoidance, reframing recovery anorexia as a targeted gut-brain protective response.
Taste preference itself proved sensitive to neurosteroid-modulated inhibition. In the gustatory insular cortex, δ-subunit containing GABAA (δGABAA) receptors mediate tonic inhibition (a continuous inhibitory signal that sets baseline neuronal excitability) and this inhibition is enhanced by the neurosteroid allopregnanolone (ALLO) (8). By mapping distribution with our Anti-GABA(A) δ Receptor (extracellular) Antibody (#AGA-014) researchers showed widespread δGABAA receptor expression. Moreover, systemic ALLO increased tonic inhibition, raised sucrose detection thresholds, and reduced sweet preference, linking hormonal state to hedonic taste processing via a defined receptor population.
In contrast, nicotine intake proved more resistant to simple pharmacological manipulation. In an operant self-administration setup, rats were trained under a fixed-ratio schedule to self-administer nicotine. During the maintenance phase, they received daily NBQX (#N-185) (5 mg/kg), an AMPA/kainate receptor antagonist, for 10 days. This regimen did not alter nicotine intake (9).
After two weeks of forced abstinence, animals were treated with NBQX or CNQX (5–6 mg/kg IV) during the last three days and immediately before reinstatement testing. Neither drug changed the total number of operant responses in the reinstatement session, but both reduced early active nose-pokes without altering the total response. AMPA/kainate antagonism therefore blunted initial drug-seeking but did not alter relapse trajectory.
Plasticity, Development, and Measurement: Making Sense of Circuits
Several studies focused on controlling or measuring circuit dynamics over time. In hippocampal slice cultures, chronic optogenetic stimulation of dentate granule cells – calibrated to physiological firing patterns – prevented denervation-induced spine loss (10). Pharmacological controls using CNQX (#C-140), D-AP5 (#D-145), and Gabazine (#G-215) ensured that light-evoked activity (and not recurrent excitation) drove the structural rescue (Figure 2). Channelrhodopsin-2 (ChR2)-expressing granule cells were repeatedly illuminated in the incubator. However, they were only illuminated after whole-cell recordings and multicell calcium imaging had been used to determine the light intensity and pulse duration required to evoke action potentials. The firing pattern of these action potentials resembled physiological granule cell bursting.
Under these calibrated conditions, chronic optogenetic stimulation preserved dendritic spine density in denervated granule cells compared to unstimulated denervated controls. Importantly, ChR2+ cells were otherwise electrophysiologically similar to non-transduced neurons, and phototoxic damage was minimized by using longer-wavelength light and antioxidant supplementation. This provided a controlled, non-invasive method to both manipulate and observe long-term circuit remodeling in vitro, anchored to real electrophysiological thresholds rather than arbitrary light protocols.
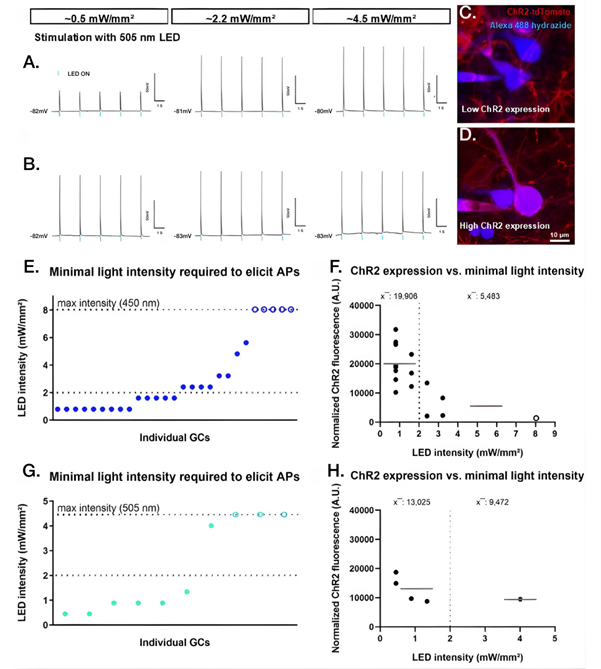
Figure 2. Neuronal responses to optogenetic stimulation depend on both light intensity and channelrhodopsin expression levels. Hippocampal dentate granule cells expressing the light-sensitive ion channel channelrhodopsin-2 (ChR2) were recorded using whole-cell patch-clamp electrophysiology while illuminated with brief pulses of blue or green light. A,B) Representative voltage recordings show differences between neurons only emerge at low light intensity (~0.5 mW/mm²): some cells spike (lower panel) while others remain at subthreshold levels (upper panel). Higher light levels drive similar firing in all neurons. C,D) Visual explanation of the different responses observed in panel A by revealing cell-to-cell differences in ChR2 expression. Neurons with higher ChR2 levels, visualized by strong tdTomato fluorescence and identified by Alexa Fluor 488-conjugated filling during recording, respond more robustly to weak light, providing a structural basis for the distinct physiological responses seen at low stimulation intensity. E) Across cells, the minimum light intensity required to trigger action potentials varied substantially, with some neurons responding to very low intensities and others requiring stronger illumination, while others failed to respond. AP- Action Potential; GC- Granule Cell. F) Neurons activated by lower light intensities showed higher levels of ChR2 expression than less responsive cells. A.U- arbitrary units. G,H) Similar relationships were observed using green (505 nm) light, with most neurons responding at low intensities when ChR2 expression was high. To ensure that the observed responses were driven directly by light activation of ChR2 rather than by network-driven recurrent excitation, experiments included pharmacological controls blocking excitatory and inhibitory synaptic transmission (CNQX, D-AP5, and gabazine). Together, these data demonstrated that effective optogenetic activation depends on both controlled light delivery and cell-specific ChR2 expression levels. Data points represent individual cells; bars indicate mean values.
Image adapted from Hanauske et al. (2025), https://doi.org/10.1038/s41598-025-88536-w. Licensed under Creative Commons Attribution 4.0 International (CC BY 4.0).
At the systems level, dopaminergic neuron development was mapped using a multi-omics approach anchored by a single surface transporter (11). Using our Anti-Dopamine Transporter (DAT) (extracellular)-FITC Antibody (#AMT-003-F) to sort dopamine transporter-positive (DAT+) neurons, researchers profiled cells from the nucleus accumbens (NAc), substantia nigra (SNc), and ventral tegmental area (VTA) at postnatal days 7, 30, and 60. They applied ultrasensitive trace proteomics and SMART-seq2 to capture region- and age-specific proteomic and transcriptomic signatures. The data showed distinct molecular profiles between NAc, SNc, and VTA DAT+ neurons that evolved over time, reflecting their divergent functional roles in reward, motor control, and other behaviors.
For sensory neurons, a STAR Protocol detailed immunopanning-based isolation of embryonic dorsal root ganglia (DRG) neurons (12). These cultures, largely free of glia, enabled combined RNAscope and immunofluorescence to profile voltage-gated sodium (NaV) channel subtypes using antibodies available in our TTX-Sensitive NaV Channel Antibody Explorer Kit (#AK-224) and TTX-Resistant NaV Channel Antibody Explorer Kit (#AK-223).
Just as circuit plasticity unfolds over time, our ability to interpret it depends on how faithfully its molecular signals can be measured. Investigators assessed how reliably brain-derived neurotrophic factor (BDNF) isoforms can be measured in human serum (13). Testing popular ELISAs against validated standards, including our human proBDNF-Biotin (#B-256-B), showed robust specificity for total BDNF and pro-BDNF but poor discrimination for mature BDNF (mBDNF). Calculating mBDNF as total BDNF minus pro-BDNF improved agreement to within ±15%. The conclusion was clear: current assays support total and pro-BDNF measurement, but mBDNF quantification remains technically limited.
Where Fate is Decided – at the Cell Surface
Across oncology, cardiology, epithelial turnover, osmosensation, feeding, addiction, and circuit plasticity, a consistent pattern emerges:
- Targeting transmembrane helices can disrupt receptor-cytoskeleton coupling and block metastasis.
- Ion channels acting as energy and pressure sensors determine extrusion, perfusion, and inflammatory escalation.
- Peripheral membrane sensors translate metabolic and hormonal states into nutrient-specific behaviors.
- Antibodies against p75NTR, TRPA1, TRPM4/7, TRPV1, DAT, δGABAA, NaV channels, and LRRC8A make these mechanisms visible and testable.
- Pharmacological tools targeting membrane proteins – including BK channel agonists (NS-1619, NS19504), AMPA/kainate receptor antagonists (NBQX, CNQX), and channel blockers such as paxilline – move studies beyond correlation, directly linking membrane signaling events to physiological outcomes.
- Reagents such as (1) Anti-DAT-FITC antibody used in specific neuronal cell sorting, (2) anti-Nav channel subtypes used in immunopanning combined with RNAscope profiling, and (3) proBDNF protein used as a standard to validate human serum results, showed the essential role of quality reagents in developing tools for answering complex biological questions.
Rather than a single breakthrough, 2025 marked a shift in emphasis. Membrane proteins are no longer passive conduits; they integrate force, energy state, and ligand context into discrete biological decisions that can now be observed, quantified, and – increasingly – pharmacologically steered. As researchers continue to address questions of force, metabolism, and signaling at the cell surface, our focus remains on developing and validating the reagents that make such work technically possible.




 Antibody")





 (extracellular) Antibody")
 Antibody")
 Antibody")
 Antibody")
 Antibody")
 δ Receptor (extracellular) Antibody")




 (extracellular)-FITC Antibody")
